LOXHD1 SNPs: Difference between revisions
Tomemerald (talk | contribs) |
Tomemerald (talk | contribs) mNo edit summary |
||
| (62 intermediate revisions by the same user not shown) | |||
| Line 1: | Line 1: | ||
'''See also:''' [[USH2A_SNPs|Usher: USH2A]] | [[CDH23_SNPs|Usher: CDH23]] | [[Opsin_evolution:_RBP3_%28IRBP%29|RBP3 (IRBP)]] | [[Opsin_evolution:_RPE65|RPE65]] | [[Opsin_evolution:_transducins|Transducins]] | |||
=== Introduction === | |||
== | [[Image:LoxConcat.jpg|left]][[Image:Loxhd1OHC.jpg|right]] | ||
LOXHD1, a large mis-annotated autosomal gene consisting solely of 16 concatenated 120-residue PLAT domains (named for <font color="magenta">p</font>olycystin/<font color="magenta">l</font>ipoxygenase/<font color="magenta">a</font>lpha-<font color="magenta">t</font>oxins), recently [http://www.ncbi.nlm.nih.gov/pubmed/19732867? surfaced] as a causative gene for age-progressive postlingual non-syndromic hearing loss type DFNB77. Of nearly a hundred known deafness genes, only MYO3 (myosin IIIA) and DFNB59 (protein product pejvakin -- echo in Persian) share this autosomal recessive progressive phenotype. | |||
Howver there is no evidence to suggest that the three encoded proteins physically interact within auditory hair cells nor that heterozygotic combinations give rise to digenic disease (as argued in Usher Syndrome). Further, with larger numbers of pedigrees, each might exhibit syndromic effects (eg affect vision or kidney function). | |||
Mouse knockouts have not been studied; delicate auditory hair cells may just be exquisitely sensitive detectors of subtle disruptions of gene function. While LOXDH1 is specific to hair cell stereocilia within the cochlea, GenBank transcripts also imply expression somewhere within testis, placenta, hippocampus, cerebellum cortex, medulla, stomach, liver, lung, and bone marrow. The protein has not been ultrastructurally localized in any of these other tissues. | |||
[[Image:LoxHippocampus.jpg|center]] | |||
Some of these transcripts are exon-skipping, for example AK302484 disables the 12th PLAT domain though retains downstream reading frame. It's unclear whether such transcripts are functional or just splicing error inevitable in a giant protein (41 exons, 2273 amino acids) and merely destined for immediate turnover. The skipped exon, LGHDGASPESCWLVEELCLAVPTQGTKYML NCNCWLAKDRGDGITSRVFDLLDAMVVNIGVK, is still 72% conserved from human to shark; nothing other than functionality could explain such conservation. | |||
Mutations do not surface during cochlear development but rather in mature stereocilia, which degenerate over time. LOXDH1 binds to the inner plasma membrane of stereocilia, perhaps organizing protein binding there or stabilizing some aspect of the membrane. As with other concatenated domain proteins (see [[CDH23_SNPs|CDC23]] and [[USH2A_SNPs|USH2A]], subtle dysfunction in one of many internally repeated domains surprisingly cannot be compensated for by the remaining intact domains. | |||
Here the mouse I1342N disrupts the hydrophobic pocket between two beta sheets of the tenth PLAT domain but the protein is still made and localized properly. The first morphological change is observed at mouse day 21: fused stereocilia and membrane ruffling at the apical hair cell surface in the basal cochlear turn. Secondary effects arise later and include damage to spiral ganglion neurons. | |||
Human allele R670* would eliminate all but the first five PLAT domains (of 16 total), possibly producing a normal but shortened protein, but might not produce protein at all due to nonsense-mediated decay. These individuals do not experience tinnitus, balance problems or vertigo, suggesting normal vestibular function even though LOXHD1 is expressed there too. | |||
One difference between cochlear and vestibular hair cells is retention of the apical kinocilium in the latter. This is lost in developing cochlear hair cells as stereocilia mature. This correlates with peak LOXHD1 expression, suggesting to [http://www.ncbi.nlm.nih.gov/pubmed/19732867? Grillet et al] that LOXDH1 might help stabilize the stereociliary bundle in the absence of kinocilium. | |||
Since retinal receptor cells are modified cilia and LOXDH1 | Recall the kinocilium is a non-motile cilium-like structure present in the crista ampullaris of the semicircular ducts and the sensory maculae of the utricle and saccule but not in the cochlear duct organ of Corti (where in mammals it degenerates following maturation of the stereociliary bundle). Since retinal receptor cells are modified cilia and LOXDH1 functions most critically in microvillar hair cells after the kinocilium is gone, a syndromic effect extending to vision would not arise based on kinocilium/photoreceptor cilia homologization. | ||
<br clear="all"> | |||
=== Phylogenetic occurence === | === Phylogenetic occurence === | ||
The PLAT domain is quite ancient, readily tracing back to prokaryotes. However in other proteins it occurs only as single copy, often in conjunction with one or more catalytic domains. A common denominator | The PLAT domain is quite ancient, readily tracing back to prokaryotes. However in other proteins it occurs only as single copy, often in conjunction with one or more catalytic domains. A common denominator in PLAT domain proteins is their association with lipid or membrane metabolism. This is perhaps mediated by extruded tryptophans which extend out from the standard beta barrel surface. | ||
LOXHD1 itself has highly conserved orthologs throughout vertebrates, indeed deuterostomes (though the Branchiostoma gene model XM_002606291 contains other domains). These species all contain mechanosensory cells with ciliary F-actin based protrusions though expression has not be localized outside of mammals. No counterparts of the gene can be found in Ecdysozoa (strictly non-cilary mechanosensation) yet lophotrochozoa (Schistosoma and likely Aplysia) have full length counterparts, as can cnidrians (Hydra, Nematostella) though gene models at this time are fragmentary. | |||
This establishes gene loss in the ancestral ecdysozoan, rather than gene innovation in Bilatera. However the overall impression is phylogenetically spotty occurence outside of deuterostomes. It's unclear whether some other gene product can compensate for lost LOXHD1 or whether it simply does not play an essential role. | |||
Most surprisingly, full length orthologs of LOXHD1 with 17 PLAT domains and very similar intronation patterns are readily recoverable from early eukaryote genomes, notably Trichoplax and the choanflagellate Monosiga. This seems to rule out an obligatory association with F-actin-based mechanosensory organelles as the basal metazoan Trichoplax exhibits only 4 distinct cell types and the early animal Monosiga is unicellular. | |||
Thus LOXDH1 is not even a metazoan innovation. Lateral gene transfer makes no sense in this context and is contradicted by the gene tree. The extraordinary conservation Monosiga to human -- surely in the 95th percentile of all genes -- establishes a long-selected intracellular role. | |||
In the plant world, occurence is restricted to Chlamydomonas and Micromonas proteins (XM_001703369, XM_002502455) with 20 PLAT domains and numerous WD40 domains (ie not necessarily orthologous to animal pure poly-PLAT proteins). A curious note on the Chlamydomonas genbank entry states "identified by comparative genomics as being present only in organisms having motile cilia." Vertebrate kinocillia are non-motile. | |||
Within deuterostomes, the large length of the gene does not sit well with gappy, incomplete and sometimes chaotic assemblies. Chicken, lamprey, amphioxus, tunicate and sea urchin are among the many that do not yield satisfactory full length gene models, in part because transcript support is limited to small regions of large proteins. However these species do contain long poly-PLAT proteins orthologous to human LOXHD1 (based on consecutive domains having consecutive best-blast). | |||
Some of these unreliable sequences are provided in the reference sequence compilation below. Ironically, the obscure Monosiga and Trichoplax have better assemblies and more accurate gene models presumably because their genomes are much smaller. | |||
Thus within vertebrates it is only feasible to compare selected PLAT motifs. Here there is a risk of using best-blast because significant cross-over matches will occur to other PLAT domains within the same protein, so some additional signature must be used for validation. | |||
=== PLAT domain structure === | |||
The structure of three proteins containing single PLAT domains have been determined. The best blastp of human LOXHD1 to these is a coral lipoxygenase/allene oxidase natural fusion protein with PDB accession [http://www.jbc.org/cgi/pmidlookup?view=long&pmid=16162493 3DY5]. Lower quality but still useful matches occur with RAB6 (accession [http://www.ncbi.nlm.nih.gov/pubmed/19141279 3CWZ]) and rabbit reticulocyte lipoxygenase (accession [http://www.ncbi.nlm.nih.gov/pubmed/9406550 2P0MA]). | |||
The basic structure of a PLAT domain consists of a sandwich of two 2-stranded beta sheets (beta barrel). The lipoxygenases are calcium-binding via extended pairs of acid residues and have conserved hydrophobic residues likely inserting into the plasma membrane but neither these nor the hydrophobic residues are conserved in LOXHD1. | |||
The independently folding PLAT domain is sometimes denoted as LH2 or [http://www.jbc.org/cgi/pmidlookup?view=long&pmid=9632630 'C2-like'] depending on the structural publication and domain tool. This introduces confusion because SMART/Pfam/InterPro utilize very distinct HMM models for C2 and PLAT/LH2. | |||
The secondary structure here is so generic that convergent evolution (non-homologous origins) must be considered, as with TIM proteins. Here however significant blastp matches at the primary sequence level unify PLAT and LH2 sequences to the exclusion of the entire C2 family. This agreement cannot have originated repeatedly by chance given the vast combinatorial space available to amino acid sequences. | |||
Further, all PLAT/LH2 domains share characteristic conserved residues. Thus PLAT and LH2 domains are in fact all homologous (descended from a single source) and so multiplicity of domain names is inappropriate. PLAT is used here rather than LH2 (<font color ="red">l</font>ipoxygenase <font color ="red">h</font>omology) because the former is a more comprehensive acronym of protein classes containing the domain (lacking only lipases). | |||
[http://www.jbc.org/content/273/26/15879.long#ref-2 C2 domains] bear an uncertain relationship to PLAT domains because a resemblance occurs only at the secondary and tertiary structural levels. The lack of convincing blastp of HMM profile matches could be due either to extreme divergence of primary sequence or to convergent evolution resulting in the same basic fold (and perhaps a similarly convergent hydrophobic intermediating function). If ever homologous, PLAT and C2 diverged in prokaryotic times. | |||
The name C2 implies calcium binding yet even here some C2 domains function independently of calcium. It isnot clear whether calcium binding is ancestral or derived. If ancestral, it has not been resolved whether binding has been lost once or multiple times in different regions of the domain tree. While PLAT domains can be structurally aligned to C2 domains, that is potentially misleading and unnecessary given three determined structures for bona fide PLAT domains. | |||
LOXHD1, which tracks back to the earliest unicellular animals, lacks capacity for calcium binding in all 16 of its domains in all descendant clades based on non-conservation of acidic liganding residues relative to coral lipoxygenase, human LOX5 and clostridial alpha-toxin PLAT domains. | |||
[[Image:AlleneLox.jpg]] | |||
=== PLAT domain evolutionary history === | |||
[[Image:LoxDomainTree.jpg|left]] | |||
The human genome contains 20 other proteins with significant blastp matching to the PLAT domain of LOXHD1. These are shown below along with their flanking domains (GPS, RUN etc). After adjusting for gene expansions (paralogy), only 4 classes occur: polycystins, lipoxygenases, lipases, and RAB6-interacting. These all contain a single PLAT domain. The final column shows its best-blastp to LOXHD1; no strong pattern emerges though perhaps PLAT:09 may conserve the most ancestral features and PLAT:15 the least. | |||
The first two columns at bottom shows the blastp score of a given human LOXHD1 PLAT domain averaged over it match to the 12 other genes; the second two columns show the average match of other genes over the 16 LOXHD1 PLAT domains. | |||
The domain tree thus looks like ((((LOXHD1,RAB6Is),polycystins),lipoxygenases),lipases). A possible parsimonious evolutionary scenario postulates a single standalone PLAT domain originally serving as an auxillary protein to various enzymes with hydrophobic substrates. Later, copies fused with these catalytic domains in separate events with gain in efficiency. | |||
[[Image:PLATrates.jpg|left]] | |||
Observe the PLAT domain is N-terminal to the catalytic domain of lipoxygenases but C-terminal to it in lipases and sandwiched internally by a repeated domain in RAB6I, indicating three separate fusion events. These fusion proteins then expanded copy number to the small paralogous gene families seen today. The individual PLAT domains then evolved at different rates, though they continued to share some common constraints (beta barrel, membrane fingers). | |||
The | The standalone gene eventually expanded to a poly-PLAT protein; today the standalone domain itself is gone (no longer represented in extant species). It is not easy to discern intermediate events: each species seems to generate a unique internal domain tree (above). In other words, individual PLAT domains may have evolved at different rates in different clades, obscuring initial relationships from order of duplication. | ||
Intermediate events may have included 1 domain tandemly duplicating to 2 and then to 4, 8, and finally 16 but other scenarios involving inhomogeneous recombination (and even higher multiplicities) are equally plausible. However the number of repeats has been very stable over many billions of years of branch length; each individual repeat seems important too in view of known point mutations. | |||
[[Image:LoxTrees.jpg|center]] | |||
<font color ="blue">PKD1L1 GPS-TM-PLAT-TM chr07:47780815 polycystin 1-like 1 pdb:3CWZ PLAT:14 36% | |||
PKD1 GPS-TM-PLAT-TM chr16: 2078712 polycystin 1 pdb:3CWZ PLAT:01 38% | |||
PKD1L2 GPS-TM-PLAT-TM chr16:79691985 polycystin 1-like 2 signal pdb:3CWZ PLAT:11 35% | |||
PKD1L3 GPS-TM-PLAT-TM chr16:70556160 polycystic kidney disease pdb:3CWZ PLAT:01 40% | |||
PKDREJ GPS-TM-PLAT-TM chr22:45030224 receptor egg jelly signal pdb:3CWZ PLAT:11 32%</font> | |||
<font color ="brown">DENND5A RUN-PLAT-RUN chr11: 9116949 RAB6 interacting 1 pdb:3CWZ PLAT:09 42% | |||
DENND5B RUN-PLAT-RUN chr12:31426424 RAB6 interacting 2 pdb:3CWZ PLAT:09 37%</font> | |||
<font color ="green">ALOX5 NH2-PLAT-lipox chr10:45189635 arachidonate 5-lipoxygenase pdb:2P0M PLAT:01 37% | |||
ALOX15 PLAT-lipox chr17: 4480963 arachidonate 15-lipoxygenase pdb:2P0M PLAT:03 26% | |||
ALOX12 PLAT-lipox chr17: 6840108 arachidonate 12-lipoxygenase pdb:2P0M PLAT:09 30% | |||
ALOX15B PLAT-lipox chr17: 7888129 arachidonate 15-lipoxygenase pdb:2P0M PLAT:02 34% | |||
ALOX12B PLAT-lipox chr17: 7916679 arachidonate 12R-lipoxygenase pdb:2P0M PLAT:09 29% | |||
ALOXE3 PLAT-lipox chr17: 7939943 arachidonate lipoxygenase 3 pdb:2P0M PLAT:16 32%</font> | |||
<font color ="magenta">LIPC LIP-PLAT chr15:56511467 lipase C pdb:2P0M PLAT:11 30% | |||
LIPG LIP-PLAT chr18:45342425 endothelial lipase pdb:2P0M PLAT:09 32% | |||
LPL LIP-PLAT chr08:19840862 lipoprotein lipase pdb:2P0M PLAT:11 24% | |||
PNLIP LIP-PLAT chr10:118295418 pancreatic lipase pdb:2P0M PLAT:16 24% | |||
PNLIPRP1 LIP-PLAT chr10:118340480 pancreatic lipase-related 1 pdb:2P0M PLAT:16 27% | |||
PNLIPRP2 LIP-PLAT chr10:118370455 pancreatic lipase-related 2 pdb:2P0M PLAT:01 31% | |||
PNLIPRP3 LIP-PLAT chr10:118177414 pancreatic lipase-related 3 pdb:2P0M PLAT:02 27%</font> | |||
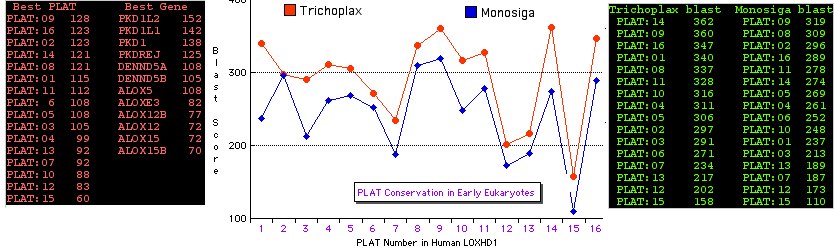
[[Image:Loxtrichmono.jpg]] | |||
Below PLAT domains | Below all PLAT domains found in human proteins are aligned with the 16 domains of human LOXHD1. The coral lipoxygenase is included because it provides the template for beta strand assignment and locating the problematic calcium binding and hydrophobic extended residues. The lower alignment is just with human LOXHD1 domains and uses an optimized seventh PLAT domain (the KE expansion is removed). | ||
[[Image:PLATalignments.jpg]] | [[Image:PLATalignments.jpg]] | ||
Individual PLAT domains | Individual PLAT domains can be quite conserved over the mammalian time scale. That is illustrated by a difference alignment of the fifth PLAT domain in 40 vertebrates (human to lamprey) below. Note columns where human sequence represents a unique change -- such as H->R in column 5 -- may be minor alleles or disease variants rather than the predominate residue in the overall human population. | ||
<pre> | <pre> | ||
01.hg18_6 MARYHVTVCTGELEGAGTDANVYLCLFGDVGDTGERLLYNCRNNTDLFEKGNADEFTIESVTMRNVRRVRIRHDGKGSGSGWYLDRVLVREEGQPESDNVEFPCLRWLDKDKDDGQLVRELLPSDSSATLK | |||
02.panTro ....R.............................................................................................................................. | |||
03.gorGor S..R...........................................................................--------------------------..................G...... | |||
04.ponAbe ....R..............T.....................................................................................-....................N.... | |||
05.rheMac ....R.........................................................................................................................N.... | |||
06.calJac ....R...........................................................K.............C...............................................N.... | |||
07.micMur ....R...........................................................M.............................................................N.... | |||
08.otoGar ....R. ............K.................-..........H.P.V.......T....................N.... | |||
09.tupBel ...R...........................................................K............A................................................N.... | |||
10.mm9_6_ ....R...........................................................K.....V.......................................................N.... | |||
11.rn4_6_ ....R...........................................................K.....V........................................Q................... | |||
12.dipOrd ....R.....................Y..................E..................K.....V.......S.S.............................................N.... | |||
13.cavPor ....R.......F.............Y...................................L.K.............................................................N.... | |||
14.speTri .V..R.....................Y......-.....-------------............K............P......E...L.....................................N.... | |||
15.oryCun ..........S.............K..........................................................TG.N.... | |||
16.ochPri ....R...........................................................K.............S....V..........................................N.... | |||
17.vicPac ...R.......V.............................I.....................K.................F.E.........................................N.... | |||
18.turTru ....R...........................................V...............K.....V...........F.......................---------...........N.... | |||
19.bosTau ....R.....................Y.....................................K.................F.E....................-....................N.... | |||
20.equCab ....R...........................................................K............A....F.E.........................................N.... | |||
21.felCat ....R...........................................................K............G....F.E.........................................N.... | |||
22.canFam ....R...........................................................K............G....F.E.........................................N.... | |||
23.myoLuc ...HR......D.R......S.....Y............T.K..DEM.Q...............K........T.V.G....F..................Y........................N.... | |||
24.pteVam ...R.........................................M.................K..........R.G....F.E.........................................N.... | |||
25.eriEur ....R.....................Y.....................................K............G....F.A..............M..........................N.... | |||
26.loxAfr ..Y.......-..M.....................A..L.K............G......E.........................................N.... | |||
27.proCap ....R.....................Y..A.......M...........R.........A..L.K............A......E..............M...---...................NN.... | |||
28.echTel ....R.....................Y..........MF.....V..............A..L.K......M.....A......-...LKK.........K..G.G....................N...R | |||
29.choHof .V..R.....................Y.........W.........M....S...Y...A..L.K.K.I.VG.....GN...F.E..............M....N...AR.Q.....I......G.N.... | |||
30.monDom ..Q.R..L...DI.....N.QAFV.....A......M.......MEICQ..........A..V.K......G.....K....F.VK..................N..F...Q..............NSR.. | |||
31.ornAna IVK.R...V..D.N......R.FI..I.........I.........T.........FV.A..LKQ......G.....GS.....AK.I.........EA.....Y.....NE....I....V.AGE.PL.. | |||
32.galGal VIK.R......MVS.S......FV..I..Q....D.V..K.I..VNK.........F..A..LKQ......G.....GS.....AK.I.........EA.....Y.....NE....I....V.AGE.PL.. | |||
33.anoCar IK.R...H..NVS.S....H.FA..I..Q....D.V.Q.SV.TVNK....S....I..A.NLKQ......G.....GS.....GK.I...D.....EAQ....Y.....NE....II...V.AGR.SL.E | |||
34.xenTro ...F.IE.K..MNK.......D.I..A..LKQ.K....G.....G.C...M.K.V...D.K..TEAI....N..F.RNE...HII...V.AGD.QN.R | |||
35.tetNig LK.RI.I...NVG.G....S.F.NII..L......SMITSK..VNK.....H...L....CLGR.S...VG...R.G.C..F..K.T.........SAI....F....HNE....I....V.AGDGRLF | |||
36.fr2_6_ LIK.R..I...NVS.G....S.F.NVI..L.......MIMSK..VNK.....H...L....SLGQ.....VG...R.G.C..F..K.M.........SAI....F....HNE....I....V.A | |||
37.gasAcu LIK.R..I...NVS.S....S.F.NVI..L.......MFMSK..VNK.....H...L....SLGQ.....VG...R.G.C..F..K.M...D.....LAT....F....RNE....I....V.AGDGRLF | |||
38.oryLat IK.R..I...TVS.S....S.F.NVI..L.......M.MSK..VNK.....H...L....WLGQ.....VG...R.G.C..F..K.M.........AAI....N....QNE....II...V.AGEG | |||
39.danRer VK.R...Y..DVS.S....H.F...I..L....D.S.I..K..INK..R......I..A.SIGP......G...R.G.C.....K.TI.....A..QA.....S..F.RNE....I....Q.FADGRLY | |||
40.petMar VK.L..IH..KHG.G......FV..Y.EQ....D.M..TS...VNK..R..V...V..C..LKR.T....G...R.GS...F..KIM.K-D.RRQCEE.....N....D.E....I....VAGGCQMLK | |||
Consensus ...yrvt..tg...g.gtda.v..cl.gd.gdtger.ly.crnn...fekgna...t..s.t$rk.r...!r...k.ggs..y$d.!lvre#.qpes.nv...c.r.l.k#k....l!...l.sgsnatlk | |||
</pre> | </pre> | ||
=== Expansion of the PLAT:07 domain === | |||
The seventh PLAT domain (human numbering) is unique domains in having a large compositionally simple expansion in the middle of the domain. It is a unique signature of the LOXHD1 orthology class that would distinguish it from other poly-PLAT domain concatenates of independent origin. That issue is completely hypothetical because no other protein in any vertebrate genome contains more than a single PLAT domain. Indeed, the LOXHD1 gene is single-copy in all species that retain it (including teleost fish). Thus even with very ancient eukaryotic divergencs, the corresponding genes can be taken as straightforward human orthologs. | |||
Presumably a loop out of the beta barrel, the expansion consists of an extremely high AG content at the dna level and encodes charged residues, primarily lysine K and glutamate E residues. These are asymmetrically distributed with the lysines N-terminal and the glutamates distal. The region will be called the KE loop of PLAT:07 here. | |||
Despite minimal net charge, this region may not be able to interact with (hydrophobic) membranes. Yet the split PLAT domain retains the potential to form a standard beta barrel. The seventh PLAT domain is quite variable even within mammals and must be hand-annotated to ensure accuracy (because its quasi-repetitive nature wreaks havoc with alignment). This section uses comparative genomics within eukaryotes to answer the following questions: | |||
* when did the high GA section arise in phylogenetic time? [early eukaryotes] | |||
* what is its source? [internal expansion, replication slippage] | |||
* did it arise from intron or retroposon retention? [species lacking the repeat also have a single exon] | |||
* can the boundaries of 7th PLAT domain be better located? [conservation on both sides of the KE region allow definition of the PLAT domain segments] | |||
* is the 44-species alignment accurate in this region? [corrections are needed when longer than human, eg platypus] | |||
* does the KE domain disrupts PLAT function? [evidently not: the split domain is highly conserved and can form a normal beta barrel] | |||
* is it a mammalian innovation or an unwanted intrusion? [this ancient expansion has variable expansion and contraction in different clades] | |||
homSap LEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLRQLLKKERLKAKLQRKKKKRKGSDEEDEGEEEESSSSEESSSEEEEMEEEEEEEE FGPGMQEVIEQHKFEAHRWLARGKEDNELVVELVPAGKPGPE 1 | |||
ornAna LEATDVGEIYKVRLGHSGEGFGSGWFIESLVLKRLVLKEVEPNPEEEKRKAKERERAREQRRKERLKAKQQRKKKKKMKKSSDDEDSEAEDSEEEEGSSEEESSSSSSEEEVEEEFGPGIKEVIDVYKFEAHRWLARDEDDKELIVELEPANRPGPE 1 | |||
galGal VEAADVGKIYKIRIGHDGKGIGDGWFLESVTLKRLATKMDETD KKKKKKKKKSEEEEEEEE TKVEEVMDVYTFVAHRWLAKDEGDKELVVELVPDGESELE 1 | |||
taeGut REAADVGKIYKIRIGHDGTGIGDGWFLESVTLKRLATKTEGSD KKKKKKKKSEEEETKE EEGMDVYTFVAHRWLAKDEGDKELVVELVPDGESDLE 1 | |||
anoCar VEAVDVGKVYKIRIGHDGKGFGDGWFLDSVVVKKLPTKVP KKKKKKKKKKTPEEEEAEE GPGIMEVYNFTPCRWLASDEEDKELVVELVPDEGSELE 1 | |||
oryLat IEALDVGKIYKIRIYHDGSGIGDGWFLETVDIKRLTMALVQVEV KKEEAPKKDKKKDKKKKKKEEEEVEIIEE MQEVVETFTFTCNRWLARDEEDGEIVVELLTEENEDLE 1 | |||
gasAcu IEAKDVGKIFKIRIGHDGSGIGSGWFLETVDVKRLILALVPKEK KKEDKKKKKKKKEDVDEEGGEE MQEVVLTYSFPCSRWLAGGEEDGELVVELLPDDAKELE 1 | |||
takRub IEAKDVGKIFKIRIGHDGLGIGSGWFLEKVYVKHLIMALVPREN KKDDKKKKKKKKKDKEDEEEVGGEE MQEVVVTYHFPCSRWLASGEDDDDLVVELLPEDAEELE 1 | |||
petMar VEAMDVGKVVKLRVGHDNSGMGSGWFLDSIVIRRLRQSSPHRPQPVDA EEDEDEEDDEEAEDED VQTYTFPCKRWLARDEDDGEIVRELLPQDCAEME 1 | |||
sacKow IEAADVAMLTKIRIGHDNSGRSAGWYLERVIIERFPPKRKM KRKRSGTPRRRGEYDEEDYDD IPETNVVNFVCNRWFAKDEEDHQIVRELLPTDEEALKGH 1 | |||
triAdh VECEDVGKLRKLRIGHDSAGMGSAWFLDKVYVRRLPPKSGKKSKET DEREEETAKKDADEPEKLDE NNYLFVANRWLSKEEGDRQTVIEISPVGVDGALA 1 | |||
monBre LACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQAR IYTFEHNDWLRKGTKAKPFMVELPLRRIETVDDN 1 | |||
=== Introns relative to PLAT domains === | |||
Here it emerges that PLAT domains correspond to 2-3 consecutive exons with conserved phasing, allowing for potential alternative splicing that reduced domain count while not introducing premature stop codons. This may reflect the origin of the 16 PLAT domains by iterated inhomogenous recombination. (to be continued). | |||
=== Reference sequences (intronated) === | === Reference sequences (intronated) === | ||
The sequences below are broken into individual exons with reading phase ([[Opsin_evolution:_ancestral_introns#Intron_location_and_phase_for_dummies|codon overhang]]) indicated by 012. Alternating colors show odd and even numbered PLAT domains. The 7th domain (which contains the charged KE expansion loop in mammals) is shown in red. | The sequences below are broken into individual exons with reading phase ([[Opsin_evolution:_ancestral_introns#Intron_location_and_phase_for_dummies|codon overhang]]) indicated by 012. Alternating colors show odd and even numbered PLAT domains. The 7th domain (which contains the charged KE expansion loop in mammals) is shown in red. In some cases, the PLAT domains are numbered and exon phases are not indicated. In the case of poor quality but phylogenetically important sequences, an approximate sequence is shown. | ||
>LOXHD1_homSap_full 41 exons|2273 aa|chr18|span 179,559 bp|no signal peptide|no conserved cysteines|16 PLAT domains|EIW --> WNW mouse deafness | >LOXHD1_homSap_full 41 exons|2273 aa|chr18|span 179,559 bp|no signal peptide|no conserved cysteines|16 PLAT domains|EIW --> WNW mouse deafness | ||
| Line 134: | Line 227: | ||
1 GTTDTFEFDSIYLGDIASLCVGHLAREDRFIPKRELAWHVKTITITEMEYGNV 2 | 1 GTTDTFEFDSIYLGDIASLCVGHLAREDRFIPKRELAWHVKTITITEMEYGNV 2 | ||
1 YFFNCDCLIPLKRKRKYFKVFEVTKTTESFASKVQSLVPVKYEVIVTTGYEPGAGTDANVFVTIFGANGDTGKRELKQKMRNLFERGSTDRFFLETLELGELRKVRLEHDSSGYCSGWLVEKVEVTNTSTGVATIFNCGRWLDKKRGDGLTWRDLFPSV* 0 | 1 YFFNCDCLIPLKRKRKYFKVFEVTKTTESFASKVQSLVPVKYEVIVTTGYEPGAGTDANVFVTIFGANGDTGKRELKQKMRNLFERGSTDRFFLETLELGELRKVRLEHDSSGYCSGWLVEKVEVTNTSTGVATIFNCGRWLDKKRGDGLTWRDLFPSV* 0 | ||
>Gallus gallus XM_425221 | >Gallus gallus gappy genome,flawed XM_425221 15 PLAT domains | ||
MKETKKNKAEEEEEEEEEEEEENEEVVEDVVEEAVEEDGEDESQQKKSKKKSKSEDSEDDGEVKKKKKKKKKTKRKEYSSEEEDDYERRKKKKKKGKKSK | |||
EKKKKKGDKSKKGKKEDNFLELYEQELRDYHSDSSNATEDEYNKKKVYEVVTVTGDVRGAGTDANVFVTLFGEFGITPKTHLTSKSSTAFERSKTDVFRV | |||
KTNNVGQIKKIRIEHDNTGLNAGWFLDRVIVTDMNRPHLRFYFPCNNWLSKEDGDGLYVRDLIGSLNPMDVPKINKYVVRVFTGEVSGSGTDADVFINIF | |||
GEKGDTGVRKLDNDKDNFEKGAEDKFTLDAPNLGRLRKINIGHNNKGGSAGWFLAKVIIEDIGNKCVYQFPVGRWFALDEDDGKIQRDILVGGTEATGIV | |||
YNVAVVTGDIRGAGTNSKIHVILHGSKGLKNSGTIFLEGGEFERARTDLFNVEIASLLSPLSRVTIGHDNCGVSSGWYCEKVVVYCPFTGIEQTFPCGKW | |||
LDEDEGDGLIERELYEMVSLRQRRLKKNPWSLWIWTSDIKNAGTDATIFFQIYGDKGKSDEMKLDNNSDNFEAGQTDKFMIELPDLGTFYKLRIWHEKRN | |||
PFAGWHLDKVTLLKTLTKDKYSFNCGRWLDINEDDNEIVRELPAEGSLVTEVMPVIKYRVTVCTGMVSGSGTDANVFVCLIGDQGDTGDRVLYKCINNVN | |||
KFEKGNADEFFVEAVTLKQVRRVRIGHDGKGGSSGWYLAKVIVREEGQPESEAVEFPCYRWLDKNEDDGQIVRELVPAGESPLLKNVSYHISVKTGDIPG | |||
ASSDSKVFIKLYGEKADTSKETLLVSDNDLGNYFERGRVDEFTIDTMDIGKINRILIGHDNVGLRSGWFLASVQITVPVQGRQYMFPCNRWLDKDEADGR | |||
VEVEVYPSEILPIEKLINYEVSVVTGDVRAAGTNAKVFMQIYGETGKTELIILENRSNNFERGATDIFEVEAADVGKIYKIRIGHDGKGIGDGWFLESVT | |||
LKRLATKMDETDKKKKKKKKKSEEEEEEEETKVEEVMDVYTFVAHRWLAKDEGDKELVVELVPDGESELEENTYEVHVLTGSVWGSGTDANVFLSIYGIE | |||
RGDTGERQLKRSNNLNKFEKGQVDVFTIKAIDLGELKKLRIRHDNSGSSPSWFLERVEIVDLKESTTYYFPCQRWLAVEEDDGQIVRELVPVDEAFVKKD | |||
SENDGQSLATLGLEQKAKSTTYIVKVKTGDKKNAGTDANVFITLYGSKDDTGIVSLKASKLNKNKFERGKIDEFTVESVDIGDLKKIKIGHDNAGNSNGW | |||
FLEWVEIDAPSLGQCLKFPCGRWLDKSEDDGAIERFIFPAELQTTEYIPFVPYEITVYTSDIFGAGTDADVFIVLYGSDGICTQQKSLCLNKREQRMYFE | |||
RNSVNQFIVELEDVGDIIEKIRIGHNGGGLNSGWHLDRVAIRRLLPNGKGSETITFPCERWLAKSEDDGEIIRELVPSDIFTEKLMKDGTLKQIEEEVED | |||
PLEVHTYKISVFTGDIYGAGTDANVFLNIYGDLGDTGERKLSKSETNFNKFERGQSYDGERRSLSESSVSLSSLDSSGNPKEKKSRLVKSAEEGLLIPYH | |||
ITVTTGTEYDSSTDSRVFIIIMGPQKVRTERLWLDLPEGKDEFADGSVEKFSVWGLDVGEIKKVEVGHDGATPESCWLMEELTIVVPTKGVMYNFVCKCW | |||
LARDKGDGLTSRILNILDADCVNVGIKILYEVTVVTGDIESGGTDAGIFMTVFGSNGNTEEMQLDKNGDRFERGQEDSFIMEIADIAPLRKMRIRTDAKG | |||
TRPDWFLERIVMRNLTNQEVATFTYGDWLSKVKNAKGSLVCEMPAMVNDEQMMEDTTYTIQVKTSDIGGAGTDANVSLILFGENGDSGTLALKESNKSNK | |||
FERNQMDEFNFPNMLSLGDLCKVRIWHDNKAYEIVTVTSNREDAETKENIWIILEGKLGRSKEFLMENSSKKRRFERRGSTDTFQFSSKNLGDIAAICVG | |||
HCPKDGKKSSAKADVYWHVKEIIITEMELCNKYFFRCNGKIPLRYKRRDYKVFECAKVIESFASKARSLVPVKYETIVVTGFEKGAGTDANVFITIFGLN | |||
GDSGKRALKQKFRNLFERGKTNRFYLETLDMGELKKVRIEHDNSGLAPGWLVERVEITNSATGVTTIFPCGKWLDENRGDGLTWRELFPRY* | |||
>LOXHD1_cioSav Ciona savigny approx gene model 15 PLAT domains | >LOXHD1_cioSav Ciona savigny approx gene model 15 PLAT domains | ||
| Line 219: | Line 314: | ||
2 K<span style="color: #0066CC;">ITYHLKITTANVKRAGTDCSINLQIFGTNGVTNCYILEKTSNRFAQGITDNISLEMEDVGKLLKMRIGHDNQ 0 | 2 K<span style="color: #0066CC;">ITYHLKITTANVKRAGTDCSINLQIFGTNGVTNCYILEKTSNRFAQGITDNISLEMEDVGKLLKMRIGHDNQ 0 | ||
0 GKNKHWNLSCVEVTVANTNQLYRFVYDDWLSLTYGKRKSLWADLPAM</span>IGDTVQLK 1 | 0 GKNKHWNLSCVEVTVANTNQLYRFVYDDWLSLTYGKRKSLWADLPAM</span>IGDTVQLK 1 | ||
2 | 2 E<span style="color: #996633;">TCLDIFVKTGNMPASSTDANVYVQLFGEYGDSGEILLKQTVSNQKPFQNNS 0 | ||
0 IDHFKIPSILKLGNLARCRIWHDNKGSSPNWYCEWLEVKEVLIPGEKNLACNWKFAFNKWLSVSDDNKQLLRDAPCS</span>EVYMNDSKGHRTIDQESIETLLTTADSMNISDLKDNPEGKL 1 | 0 IDHFKIPSILKLGNLARCRIWHDNKGSSPNWYCEWLEVKEVLIPGEKNLACNWKFAFNKWLSVSDDNKQLLRDAPCS</span>EVYMNDSKGHRTIDQESIETLLTTADSMNISDLKDNPEGKL 1 | ||
<span style="color: #0066CC;">2 VYEVVIETGNLKDSGTTCDAWIILEGKHGRSPKLELVNQVGNPILQINQMNTFQ 2 | <span style="color: #0066CC;">2 VYEVVIETGNLKDSGTTCDAWIILEGKHGRSPKLELVNQVGNPILQINQMNTFQ 2 | ||
| Line 226: | Line 321: | ||
2 ESIQRIIIEHDNTGVSPDWYLDKVLITNQTNNQIHLFQCYQWISKKKGDCRLWKELLVS</span>N* 0 | 2 ESIQRIIIEHDNTGVSPDWYLDKVLITNQTNNQIHLFQCYQWISKKKGDCRLWKELLVS</span>N* 0 | ||
>Trichoplax adhaerens (trichoplax) Metazoa XM_002107971 17 PLAT domains [http://www.ncbi.nlm.nih.gov//genomes/geblast.cgi?bact=off&gi=6008 blast] | >LOXHD1_triAdh Trichoplax adhaerens (trichoplax) Metazoa XM_002107971 17 PLAT domains [http://www.ncbi.nlm.nih.gov//genomes/geblast.cgi?bact=off&gi=6008 blast] | ||
0 MSASLAMQELMSDPSGHFAPKPPPGRSKINRKRPQT 1 | 0 MSASLAMQELMSDPSGHFAPKPPPGRSKINRKRPQT 1 | ||
2 APGRQITTASKAPKLWRSSTYNEPNLKRRKFIVTLNHKKDISSISRNAPYANRAYTTAMATAARTYSNALNKGQQKEIIPIYNPLCDPHLNDYYARKFGLLNSRD 0 | 2 APGRQITTASKAPKLWRSSTYNEPNLKRRKFIVTLNHKKDISSISRNAPYANRAYTTAMATAARTYSNALNKGQQKEIIPIYNPLCDPHLNDYYARKFGLLNSRD 0 | ||
| Line 261: | Line 356: | ||
2 ASFRSNSTDLGEMVSASGRRASSRKGPVMAPIDEK 1 | 2 ASFRSNSTDLGEMVSASGRRASSRKGPVMAPIDEK 1 | ||
2 CVPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVS 1 | 2 CVPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVS 1 | ||
2 | 2 HNSYMPDSGIYLKEIEVDVPTIGNKYIFPCNRWLAKDKDDSKTSRIFTAANAQVTSYTKPYELTIHTGDVQNA 1 | ||
2 GTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIP 0 | 2 GTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIP 0 | ||
0 LELDDVGTIKKIRIGHDGKGSRTDWYLEK 0 | 0 LELDDVGTIKKIRIGHDGKGSRTDWYLEK 0 | ||
| Line 271: | Line 366: | ||
2 ELKPVKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* 0 | 2 ELKPVKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* 0 | ||
> | <span style="color: #0066CC;">>LOXHD1_triAdh Trichoplax adhaerens (trichoplax) Metazoa XM_002107971 17 PLAT domains [http://www.ncbi.nlm.nih.gov//genomes/geblast.cgi?bact=off&gi=6008 blast] | ||
MSASLAMQELMSDPSGHFAPKPPPGRSKINRKRPQT<span style="color: #996633;">APGRQITTASKAPKLWRSSTYNEPNLKRRKFIVTLNHKKDISSISRNAPYANRAYTTAMATAARTYSNALNKGQQKEIIPIYNPLCDPHLNDYYARKFGLLNSRD</span>ESRKNRKQ<span style="color: #996633;">AG | |||
01 VAYQFGVKTGDKKGSDTDA</span>VYIQVIGTKDKIPKKRLFKKQETEKTERGNLFKFDKSTVEKFAVQHRDIGDPVKLIVE<span style="color: #996633;">HDGNEKRHGWFLEEITLTNIQSKKSWLFPCHKWLSKYEGDRKLCYE LKPLAKAGK | |||
02 A</span>VYEVSVLTGDKRGAGTDANVSVTLFGKHTSSPKIQLLKS<span style="color: #996633;">SKHKNPFERNNTDEFKIRTRDVGKLSKIRIEHDNAGFGPGWFLDK</span>VIICNLEKPNVKYYCPCNQWLAKDVGDKSISRD LTAYTDPNAAPS<span style="color: #996633;">A | |||
03 YVYIVHTFTGNKRGAGTDANVYAVIFGDSGDTGEKRLDNSKNNFEKSR</span>KDTFKLSCSCVGKLERLRIRHDNTGLFAGWYLDKV<span style="color: #996633;">VVEDPQEQQSYTFYCRRWLSKTEDDGEICRDLI VSASGDDGDDSVAPK</span>G | |||
04 YPYHIHVTTSDVKNAGTDAEVYVVMHGEGKKSKELNSGKLVLANSEKKKNTFERAMTDIFHMECAEMLSPLTKLTVGHDNKGLAAGWHLDRa<span style="color: #996633;">IVIDCPTTGIEQTFLCQQWLDRKAGDGLTERELVEA FDMRKTRRP</span>K | |||
05 QLWFAWIWTSDIRGAGTDANVSMQIYGDKGKSQEIKLGNNTDNFEQATLDKFK<span style="color: #996633;">LEIDQVGVPYKLRIGHDNSNAFPGWHLDKVKLENMNDKEQYLFNCNRWLSRSEEDNEIIRELPAS GPNCPNYP</span>I | |||
06 VIYEVSVHTGNKMGGGTDANVFIKIYGELGDSGYRPLKSSKSHNNKFERNQVDVFHIEAVTLKALKKIKIGHDGNNP<span style="color: #996633;">GAGWFLDKVVIKELNGEASNEFPCNR</span>WLSKSEDDGQIVRELFLK SDTPLLK<span style="color: #996633;">T | |||
07 TSYHISVKTGDVRNAGTDANVFIQIFGAKDDTGRVRLKQSLNTSNKFERNRIDKFIIEAAQIGK</span>IEKIIIGHDGKGLGSGWFLDYIELDVPSVGRLYRFSCHQWFDSTEGDRKVERELYPS ECIKSAA<span style="color: #996633;">K | |||
08 IPYQISVHTGDIRHAGTDSNVFAVIYGENGKTEELKLRNKSDNFERGQVDVFK</span>VECEDVGKLRKLRIGHDSAGMGSAWFLDK<span style="color: #996633;">VYVRRLPPKSGKKSKETDEREEETAKKDADEPEKLDENNYLFVANRWLSKEEGDRQTVIEIS PVGVDGALA</span>E | |||
09 MTYTIRVITGNKFGCGTNANVFINMYGEEGDSGERQLKKSETHTDKFERNQ<span style="color: #996633;">EDVFKISCLSLGELKKIKIRHDNSGFRPAWFLDKVIIEVGESKYQFMCDRWLAKDEDDGQISRELLPQ SDEQSRAEAIGASKDLQKK</span>VAS | |||
10 TTYNVSVTTGDIKGAGTDANVHIVLYGEKDDTGLIHLKNSTTHSNKFERNQEDRFVVEAIDIGELKKIK<span style="color: #996633;">IGHDNKGGMAGWFLNKVEIDIPSLGRRLLFPCGRWIDKGKDDGALER ELYPLNEAEETYRP</span>H | |||
11 IPYEVTVYTTDKRGASTGANVYVVIYGEENQTEQASLEPDKKRRKQYFKNGAIDKFVLE<span style="color: #996633;">LDDVGEEITKLRIGHDGKGWGAGWHLDKVEICRLLDGGKASKKFTFQCNRWLASDEDDGAIVRELVPS EIVEKSSKDGGQVKTKVTKPTDGLK</span>V | |||
12 KPYTIHVFTGDVDGAGTNANVFLTIFGESGDSGERKLAKSDTHYDKFERNQ<span style="color: #996633;">EDIFHIEAADLGRLFKVKIRHDNTGSLFSPAWFLNRIEIVDDENEETTAFPCERWLAKKKDDGKIDRTLFVK GWEGDTSSVATTRSK</span>ASFRSNSTDLGEMVSASGRRASSRKGPVMAPIDEK<span style="color: #996633;">C | |||
13 VPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVS</span>HNSYMPDSGIYLKEIEVDVPTIGNKYIFPCNRWLAKDKDDSKTSRIFTAA | |||
14 NAQVTSYTA<span style="color: #996633;">GTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIP</span>LELDDVGTIKKIRIGHDGKGSRTDWYLEK<span style="color: #996633;">ASIQRMDTLDMYMFRANQWFSKKIDDKKLVREI PAETSKEGATTIK</span>K | |||
15 INYVLSTHTSDKRGSGTDANVFVIIFGENGDSGEIALKKSETNWNKFEKGQTDVFLINDRLSLGRLQKLRIWHDNA<span style="color: #996633;">GFGASWHLASVDIVDESTGVKYTFPCDKWLSKSNGDKLILREL PCAETAGNTAASKATKQESKSGK</span> | |||
16 AEYEIAFTTGTEKRAGTNQDVAIVLKGKSEKSREFLIENNEDKKYFSKGKTNKFTYTCKPLGDITKAIVSHRESAIGEEPDSKNSSWYLKVVTVVHKASGTT<span style="color: #996633;">YKFPCNKWIDLDDDEENNSSVTLKC KSADVAASSKAKPV</span>ELKP | |||
17 VKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* </span> | |||
<span style="color: #0066CC;">>Monosiga brevicollis (monosiga) Eukaryota|Choanoflagellates XM_001742822 17 PLAT domains [http://www.ncbi.nlm.nih.gov//genomes/geblast.cgi?bact=off&gi=5881 blast] exons colored; PLAT domains numbered | |||
MADLPVPTTALQRLQLERSRYSFAAA<span style="color: #996633;">DPPPSFVQRYTSADVLSH</span>RLYAQGLASDVGPGSHSSL<span style="color: #996633;">GMSRRVELPPYNPLNDPALANYFARKFEWNSTRSVGAGSGSRRSASASLTRTRQPVRGASAHGHTKSRTKSSHRK | |||
01 GHASLEIFTSKRSNPVSKSEKYITVVGTKGRSDAIQLAAPGVHFRAGNKDVFQVNLTGIGKPT</span>KVILENTGTKRTDGWCVSKVVLVKKTDKGTRRYRFSGPVWL<span style="color: #996633;">SKHHDEMKLKR VLHIDDEDIGSG | |||
02 NG</span>YRIDCYTGDVANAGTDAVATIQLFGSKGQSPMVELRRSDGQAFQRARVATFTLD<span style="color: #996633;">DLANLGKLKKLVISHNGHGMASGWFLDK</span>IIVTSLSSNKATVFPCDAWLDRKNGRSK<span style="color: #996633;">ELVAR TAAAGAEGM | |||
03 TTFTIRVMTGDRRGAGT</span>DANVQCTLFGEDGES<span style="color: #996633;">GPHTLNTSRNDFRRGHTDVFAVSSRKIGTLKRLRIWHDNGGAGP</span>AWFLDAVEVVDEASGQTYRFECNRWLAKDEDDGQISRELTCN GDSSWGL | |||
04 KSYKLTIFTGDKRNAGTSANVFCKLVGERGASDNVILENSSKNFQRDRTDIFTVEASDLGSLRHIVLGHDNHGM<span style="color: #996633;">GAGWYVERFSLEVPSEGKLYNVDVKQW</span>FATDMSDGAIERTFRLD NAETLDIAQR | |||
05 LDWKCTIYTSDVANAGTDANVFMQV<span style="color: #996633;">YGKKGKTDVVPLKNKSDTFER</span>GQTDELRVQLINVGSLRKLRVWHDNKGMASGWHLDRIVLSRDGEEY<span style="color: #996633;">IFPCAEWLAVSEGDKEIVRELPAT GPNVKKPLQL | |||
06 VEYTVRVATGHARFAGTNADVFVMLTGELGDSGKRALLRSQTNR</span>NKFERGKEDVFTVAAVDLGKLTSVTVGHNNAGTSAGWFLDKIVVLDPRRG<span style="color: #996633;">EEEEFPCHRWLAVDADDGQIERELVPK MAEHQAAAT | |||
07 TTYIVKIKTGDVRHAGTDANV</span>FVQLFGKTGESTQLKLRNSETYSDAFERNKMDIFKFELLDLGDLSRILVGHDNKGMGAAWFLDYVEVEVPSIRTRWKFPCSRWFSKSQDDGLTER EIYAEKEAGEPMEEDVS | |||
08 APYLFRFYTSDVAFAGTDANV<span style="color: #996633;">SVVLYGDEGKTEELVVNNQSDNFERGKA</span>DDFKLACKPVGRPSKIRLSAHGGGMSADWHLEK<span style="color: #996633;">IEVHELGQARIYTFEHNDWLRKGTKAKPFMVEL PLRRIETVDDNGREVVEELALDANK | |||
09 RTYRVKVHTGDQKGAGTDAN</span>VYVNLHGSLGDSGDRHLKNSLTHT<span style="color: #996633;">NKFQRKTVDEFDIDAVTLGDINKVKVWHDNA</span>GLGAAWYLEKIEVVDTADDKTYIFPCAQWFAKSMGDGQIAREL GVLEEQ<span style="color: #996633;">KPADFQTKNVG | |||
10 FKYRISVHTSDVKHAGTDANVDIVLYGEKGDTGKIRLAKSETHRDMW</span>ERGNCDVFTVSAIELGDLKRVDIMHDGKGVGSGWHLNKVVVDAPQAGKTWTFMCDAWLDKATDDGTMAK TLYASADAIEEYSAH | |||
11 VPYEIIIKTSDVRNAGTDANVFIDLYGRDQEERDLTAHHEFKDAVKAHFERNLEDRFNVELPDVGSIYKIRLGHDGKGMSSSWHVASVVVINQRTHERFEFPCDAWLSKDKDDKKLVREFAVG EVKALEGDKVVRRESILSLQE | |||
12 AIYKIHVFTGDIKHAGTDANIYVQIFGDTGDSGEIKLEKSETYRDKFERGHEDIFTHRCLDLGPLRKIKVRSDGKGLMGGDWYLDRVEVHQENDLSEPPVRFVCQDWFKRGKQEGDTLEREITAQ | |||
VDAAVAMKEATALEDEAARLQRKATTLSRKSTKGSPLRKGTGTTETNAELEEAQRQANIARRKADEAKERAGLAGADDTQ | |||
13 LEYKVTVYTGTDTQAGTTANVWLQLFGEKDAPLTPAPPSPSKLSRSGSLFGRRRRASSDAQSVSSSLSAGASGPRETSTGRLQLNNAPADLQSGAKTTFTVTGLDVGELVGLEIGHDDERDKWYLEQVVVEVPKSATHAARRYEFKAGVWLAARADGSTSGSAK GKSKASVRLQPSEIHAGSERI | |||
14 LEYTLKVYTASADGAGCTAVPQVQLFGDKHTTEALPLRAGGDILPASVVETQHRVPDLGALLKVRLMVPRGSSWTVEKVEFGRAGQTPITFVGSDGSAVTLGAERLS FDFLPAAAPASSGGKGKRRGSTTSSVAVAQQ | |||
15 TSYRVYVTTADERGTGTDANVSIILYGAMGDSGEHSLTKSETFDDPFERGNTDVFTLEVPDLGELQRARIWHDGKGMFSSWKLDKIVVVVEATQSRYELPCGQWLSKNKGDKQLTRDLAVASKR VNALT | |||
16 GTYKLEVATDSRAGGGCKGPVRIMLLDAEKNQLPLTLEPPGGEFAPGSVEHLVFDNVMLLGPLTELRIRRKPSGASSRRGAEDDDEDDNEASGASSQSGSAVSPWHLEHIIVKHLQSGQSFVFKGPSKGLSRSRAKLSVHTEA TTEEAVAQATKASV | |||
17 AQYEVAVTTGTERGAGTDSN<span style="color: #996633;">VFVTLFGKNGDSGERALAKSKTFRNMFESGNTDVFD</span>VECQDLGELTKIEVK<span style="color: #996633;">SDLKGFGAAWQLDKIKVTRTGSQNSWQFKCDQWFDKKQGAEHTFSVA S*</span></span> | |||
>LOXHD1_homSap_full dna: each line is an exon | >LOXHD1_homSap_full dna: each line is an exon | ||
| Line 343: | Line 451: | ||
=== Reference PLAT domain sets === | === Reference PLAT domain sets === | ||
<span style="color: #990099;">>LOXHD1_homSap_PLAT:01 | |||
<span style="color: #990099;"> | |||
RVYEVVTATGDVRGAGTDANVFITLFGENGLSPKLQLTSKSKSAFEKGNVDVFRVRTNNVGLIYKVRIEHDNTGLNASWYLDHVIVTDMKRPHLRYYFNCNNWLSKVEGDRQWCRD | RVYEVVTATGDVRGAGTDANVFITLFGENGLSPKLQLTSKSKSAFEKGNVDVFRVRTNNVGLIYKVRIEHDNTGLNASWYLDHVIVTDMKRPHLRYYFNCNNWLSKVEGDRQWCRD | ||
> | >LOXHD1_homSap_PLAT:02 | ||
NKYEVKVYTGDVIGAGTDADVFINIFGEYGDTGERRLENEKDNFEKGAEDRFILDAPDLGQLMKINVGHNNKGGSAGWFLSQIVIEDIGNKRKYDFPLNRWLALDEDDGKIQRDILVGG | NKYEVKVYTGDVIGAGTDADVFINIFGEYGDTGERRLENEKDNFEKGAEDRFILDAPDLGQLMKINVGHNNKGGSAGWFLSQIVIEDIGNKRKYDFPLNRWLALDEDDGKIQRDILVGG | ||
> | >LOXHD1_homSap_PLAT:03 | ||
ITYIVTVFTGDVRGAGTKSKIYLVMYGARGNKNSGKIFLEGGVFDRGRTDIFHIELAVLLSPLSRVSVGHGNVGVNRGWFCEKVVILCPFTGIQQTFPCSNWLDEKKADGLIER | ITYIVTVFTGDVRGAGTKSKIYLVMYGARGNKNSGKIFLEGGVFDRGRTDIFHIELAVLLSPLSRVSVGHGNVGVNRGWFCEKVVILCPFTGIQQTFPCSNWLDEKKADGLIER | ||
> | >LOXHD1_homSap_PLAT:04 | ||
FPWSLWVWTTDLKKAGTNSPIFIQIYGQKGRTDEILLNPNNKWFKPGIIEKFRIELPDLGRFYKIRVWHDKRSSGSGWHLERMTLMNTLNKDKYNFNCNRWLDANEDDNEIVREM | FPWSLWVWTTDLKKAGTNSPIFIQIYGQKGRTDEILLNPNNKWFKPGIIEKFRIELPDLGRFYKIRVWHDKRSSGSGWHLERMTLMNTLNKDKYNFNCNRWLDANEDDNEIVREM | ||
> | >LOXHD1_homSap_PLAT:05 | ||
ARYHVTVCTGELEGAGTDANVYLCLFGDVGDTGERLLYNCRNNTDLFEKGNADEFTIESVTMRNVRRVRIRHDGKGSGSGWYLDRVLVREEGQPESDNVEFPCLRWLDKDKDDGQLVRELLPS | ARYHVTVCTGELEGAGTDANVYLCLFGDVGDTGERLLYNCRNNTDLFEKGNADEFTIESVTMRNVRRVRIRHDGKGSGSGWYLDRVLVREEGQPESDNVEFPCLRWLDKDKDDGQLVRELLPS | ||
> | >LOXHD1_homSap_PLAT:06 | ||
FRYHISLKTGDVSGASTDSRVYIKLYGDKSDTIKQVLLVSDNNLKDYFERGRVDEFTLETLNIGNINRLVIGHDSTGMHASWFLGSVQIRVPRQGKQYTFPANRWLDKNQADGRLEV | FRYHISLKTGDVSGASTDSRVYIKLYGDKSDTIKQVLLVSDNNLKDYFERGRVDEFTLETLNIGNINRLVIGHDSTGMHASWFLGSVQIRVPRQGKQYTFPANRWLDKNQADGRLEV | ||
> | >LOXHD1_homSap_PLAT:07 | ||
VHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQLEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLR | VHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQLEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLR | ||
> | >LOXHD1_homSap_PLAT:08 | ||
NTYEVQVVTGNVPKAGTDANVYLTIYGEEYGDTGERPLKKSDKSNKFEQGQTDTFTIYAIDLGALTKIRIRHDNTGNRAGWFLDRIDITDMNNEITYYFPCQRWLAVEEDDGQLSRELLPV | NTYEVQVVTGNVPKAGTDANVYLTIYGEEYGDTGERPLKKSDKSNKFEQGQTDTFTIYAIDLGALTKIRIRHDNTGNRAGWFLDRIDITDMNNEITYYFPCQRWLAVEEDDGQLSRELLPV | ||
> | >LOXHD1_homSap_PLAT:09 | ||
TTFSVTIKTGVKKNAGTDANVFITLFGTQDDTGMTLLKSSKTNSDKFERDSIEIFTVETLDLGDLWKVRLGHDNTGKAPGWFVDWVEVDAPSLGKCMTFPCGRWLAKNEDDGSIIRDLFHA | TTFSVTIKTGVKKNAGTDANVFITLFGTQDDTGMTLLKSSKTNSDKFERDSIEIFTVETLDLGDLWKVRLGHDNTGKAPGWFVDWVEVDAPSLGKCMTFPCGRWLAKNEDDGSIIRDLFHA | ||
> | >LOXHD1_homSap_PLAT:10 | ||
VPYEITLYTSDVFAAGTDANIFIIIYGCDAVCTQQKYLCTNKREQKQFFERKSASRFIVELEDVGEIIEKIRIGHNNTGMNPGWHCSHVDIRRLLPDKDGAETLTFPCDRWLATSEDDKKTIRELVP | VPYEITLYTSDVFAAGTDANIFIIIYGCDAVCTQQKYLCTNKREQKQFFERKSASRFIVELEDVGEIIEKIRIGHNNTGMNPGWHCSHVDIRRLLPDKDGAETLTFPCDRWLATSEDDKKTIRELVP | ||
> | >LOXHD1_homSap_PLAT:11 | ||
VLYSVQIFTGNIPGAGTDAKVYITIYGDLGDTGERYLGKSENRTNKFERGTADTFIIEAADLGVIYKIKLRHDNSKWCADWYVEKVEIWNDTNEDEFLFLCGRWLSLKKEDGRLERLFYEK | VLYSVQIFTGNIPGAGTDAKVYITIYGDLGDTGERYLGKSENRTNKFERGTADTFIIEAADLGVIYKIKLRHDNSKWCADWYVEKVEIWNDTNEDEFLFLCGRWLSLKKEDGRLERLFYEK | ||
> | >LOXHD1_homSap_PLAT:12 | ||
IPYYVSVTTGKHKDAATDSRAFIFLIGEDDERSKRIWLDYPRGKRGFSRGSVEEFYVAGLDVGIIKKIELGHDGASPESCWLVEELCLAVPTQGTKYMLNCNCWLAKDRGDGITSRVFD | IPYYVSVTTGKHKDAATDSRAFIFLIGEDDERSKRIWLDYPRGKRGFSRGSVEEFYVAGLDVGIIKKIELGHDGASPESCWLVEELCLAVPTQGTKYMLNCNCWLAKDRGDGITSRVFD | ||
> | >LOXHD1_homSap_PLAT:13 | ||
VLYEMTVWTGDVVGGGTDSNIFMTLYGINGSTEEMQLDKKKARFEREQNDTFIMEILDIAPFTKMRIRIDGLGSRPEWFLERILLKNMNTGDLTMFYYGDWLSQRKGKKTLVC | VLYEMTVWTGDVVGGGTDSNIFMTLYGINGSTEEMQLDKKKARFEREQNDTFIMEILDIAPFTKMRIRIDGLGSRPEWFLERILLKNMNTGDLTMFYYGDWLSQRKGKKTLVC | ||
> | >LOXHD1_homSap_PLAT:14 | ||
TSYTVAVKTSDILGAGTDANVFIIIFGENGDSGTLALKQSANWNKFERNNTDTFNFPDMLSLGHLCKLRVWHDNKGIFPGWHLSYVDVKDNSRDETFHFQCDCWLSKSEGDGQTVRDFAC | TSYTVAVKTSDILGAGTDANVFIIIFGENGDSGTLALKQSANWNKFERNNTDTFNFPDMLSLGHLCKLRVWHDNKGIFPGWHLSYVDVKDNSRDETFHFQCDCWLSKSEGDGQTVRDFAC | ||
> | >LOXHD1_homSap_PLAT:15 | ||
TTYEIVIETGNGGETRENVWLILEGRKNRSKEFLMENSSRQRAFRKGTTDTFEFDSIYLGDIASLCVGHLAREDRFIPKRELAWHVKTITITEMEYGNVYFFNCDCLIPLKRKRKYFKVF | TTYEIVIETGNGGETRENVWLILEGRKNRSKEFLMENSSRQRAFRKGTTDTFEFDSIYLGDIASLCVGHLAREDRFIPKRELAWHVKTITITEMEYGNVYFFNCDCLIPLKRKRKYFKVF | ||
> | >LOXHD1_homSap_PLAT:16 | ||
VKYEVIVTTGYEPGAGTDANVFVTIFGANGDTGKRELKQKMRNLFERGSTDRFFLETLELGELRKVRLEHDSSGYCSGWLVEKVEVTNTSTGVATIFNCGRWLDKKRGDGLTWRDLFPS | VKYEVIVTTGYEPGAGTDANVFVTIFGANGDTGKRELKQKMRNLFERGSTDRFFLETLELGELRKVRLEHDSSGYCSGWLVEKVEVTNTSTGVATIFNCGRWLDKKRGDGLTWRDLFPS | ||
</span> | </span><font color="green"> | ||
>PKD1L1_homSap_PLAT chr7: 47780815 polycystin-1L1 GPS-TM-LH2-TM pdb:3CWZ | |||
> | QLYAVVIDTGFRAPARLTSKVYIVLCGDNGLSETKELSCPEKPLFERNSRHTFILSAPAQLGLLRKIRLWHDSRGPSPGWFISHVMVKELHTGQGWFFPAQCWLSAGRHDGRVERELTC | ||
>PKD1_homSap_PLAT chr16: 2078712 polycystin 1 GPS-TM-LH2-TM pdb:3CWZ | |||
FKYEILVKTGWGRGSGTTAHVGIMLYGVDSRSGHRHLDGDRAFHRNSLDIFRIATPHSLGSVWKIRVWHDNKGLSPAWFLQHVIVRDLQTARSAFFLVNDWLSVETEANGGLVEK | |||
>PKD1L2_homSap_PLAT chr16: 79691985 polycystin 1-like 2 signal...GPS-TM-LH2-TM pdb:3CWZ | |||
YHYLVTVYTGHRRGAATSSKVTVTLYGLDGEREPHHLADPDTPVFERGAVDAFLLSTLFPLGELRSLRLWHDNSGDRPSWYVSRVLVYDLVMDRKWYFLCNSWLSINVGDCVLDKVFPVA | YHYLVTVYTGHRRGAATSSKVTVTLYGLDGEREPHHLADPDTPVFERGAVDAFLLSTLFPLGELRSLRLWHDNSGDRPSWYVSRVLVYDLVMDRKWYFLCNSWLSINVGDCVLDKVFPVA | ||
> | >PKD1L3_homSap chr16:70,556,160 GPS-TM-LH2-TM polycystic kidney disease 1-like 3 | ||
FHYLIQVYTGYRRSAATTAKVVITLYGSEGRSEPHHLCDPQKTVFERGGLDVFLLTTWTSLGNLHSLRLWHDNSGVSPSWYVSQVIVCDMAVKRKWHFLCNCWLAVDLGDCELDRVFIPV | |||
>PKDREJ_homSap_PLAT chr22: 45030224 receptor for egg jelly-like signal...GPS-TM-LH2-TM pdb:3CWZ | |||
CYLVTIFTGSRWGSGTRANVFVQLRGTVSTSDVHCLSHPHFTTLYRGSINTFLLTTKSDLGDIHSIRVWHNNEGRSPSWYLSRIKVENLFSRHIWLFICQKWLSVDTTLDRTFHVT | CYLVTIFTGSRWGSGTRANVFVQLRGTVSTSDVHCLSHPHFTTLYRGSINTFLLTTKSDLGDIHSIRVWHNNEGRSPSWYLSRIKVENLFSRHIWLFICQKWLSVDTTLDRTFHVT | ||
> | >DENND5A_homSap_PLAT chr11: 9116949 RAB6 interacting 1 RUN-LH2-RUN pdb:3CWZ | ||
LIHILIVPSKKLGGSMFTANPWICISGELGETQIMQIPRNVLEMTFECQNLGKLTTVQIGHDNSGLYAKWLVEYVMVRNEITGHTYKFPCGRWLGKGMDDGSLER | |||
>DENND5B_homSap_PLAT chr12: 31426424 RAB6 interacting 2 RUN-LH2-RUN pdb:3CWZ | |||
PYRSVIIPIKKLSNAIITSNPWICVSGELGDTGVMQIPKNLLEMTFECQNLGKLTTVQIGHDNSGLLAKWLVDCVMVRNEITGHTYRFPCGRWLGKGIDDGSLER | |||
>ALOX5_homSap_PLAT chr10: 45189635 arachidonate 5-lipoxygenase NH2-LH2-lipox pdb:2P0M | |||
PSYTVTVATGSQWFAGTDDYIYLSLVGSAGCSEKHLLDKPFYNDFERGAVDSYDVTVDEELGEIQLVRIEKRKYWLNDDWYLKYITLKTPHGDYIEFPCYRWITGDVEVVLRDG | |||
>ALOX15_homSap_PLAT chr17: 4480963 arachidonate 15-lipoxygenase NH2-LH2-lipox pdb:2P0M | |||
GLYRIRVSTGASLYAGSNNQVQLWLVGQHGEAALGKRLWPARGKETELKVEVPEYLGPLLFVKLRKRHLLKDDAWFCNWISVQGPGAGDEVRFPCYRWVEGNGVLSLPEG | |||
> | >ALOX15_oryCun Oryctolagus cuniculus (rabbit) arachidonate 15-lipoxygenase 15S-LOX1 PDB:P0M | ||
GVYRVCVSTGASIYAGSKNKVELWLVGQHGEVELGSCLRPTRNKEEEFKVNVSKYLGSLLFVRLRKKHFLKEDAWFCNWISVQALGAAEDKYWFPCYRWVVGDGVQSLPVG | |||
> | >ALOX12_homSap_PLAT chr17: 6840108 arachidonate 12-lipoxygenase NH2-LH2-lipox pdb:2P0M | ||
GRYRIRVATGAWLFSGSYNRVQLWLVGTRGEAELELQLRPARGEEEEFDHDVAEDLGLLQFVRLRKHHWLVDDAWFCDRITVQGPGACAEVAFPCYRWVQGEDILSLPEG | |||
> | >ALOX15B_homSap_PLAT chr17 7,888,129 arachidonate 15-lipoxygenase NH2-LH2-lipox pdb:2P0M | ||
AEFRVRVSTGEAFGAGTWDKVSVSIVGTRGESPPLPLDNLGKEFTAGAEEDFQVTLPEDVGRVLLLRVHKAPPVLPLLGPLAPDAWFCRWFQLTPPRGGHLLFPCYQWLEGAGTLVLQEG | |||
> | >ALOX12B_homSap_PLAT chr17: 7916679 arachidonate 12R-lipoxygenase NH2-LH2-lipox pdb:2P0M | ||
ATYKVRVATGTDLLSGTRDSISLTIVGTQGESHKQLLNHFGRDFATGAVGQYTVQCPQDLGELIIIRLHKERYAFFPKDPWYCNYVQICAPNGRIYHFPAYQWMDGYETLALREA | |||
> | >ALOXE3_homSap_PLAT chr17: 7939943 arachidonate lipoxygenase 3 NH2-LH2-lipox pdb:2P0M | ||
AVYRLCVTTGPYLRAGTLDNISVTLVGTCGESPKQRLDRMGRDFAPGSVQKYKVRCTAELGELLLLRVHKERYAFFRKDSWYCSRICVTEPDGSVSHFPCYQWIEGYCTVELRPG | AVYRLCVTTGPYLRAGTLDNISVTLVGTCGESPKQRLDRMGRDFAPGSVQKYKVRCTAELGELLLLRVHKERYAFFRKDSWYCSRICVTEPDGSVSHFPCYQWIEGYCTVELRPG | ||
> | >LIPC_homSap LIP-PLAT chr15:56511467 lipase C | ||
YHYQLKIQFINQTETPIQTTFTMSLLGTKEKMQKIPITLGKGIASNKTYSFLITLDVDIGELIMIKFKWENSAVWANVWDTVQTIIPWSTGPRHSGLVLKTIRVKAGETQQRMTFCSEN | |||
>LIPG_homSap LIP-PLAT chr18:45342425 endothelial lipase | |||
YHYQMKIHVFSYKNMGEIEPTFYVTLYGTNADSQTLPLEIVERIEQNATNTFLVYTEEDLGDLLKIQLTWEGASQSWYNLWKEFRSYLSQPRNPGRELNIRRIRVKSGETQRKLTFCTEDPEN | |||
>LPL_homSap LIP-PLAT chr8:19840862 lipoprotein lipase | |||
FHYQVKIHFSGTESETHTNQAFEISLYGTVAESENIPFTLPEVSTNKTYSFLIYTEVDIGELLMLKLKWKSDSYFSWSDWWSSPGFAIQKIRVKAGETQKKVIFCSREKVSHLQKGKAPAVFVKC | |||
>PNLIP_homSap LIP-PLAT chr10:118295418 pancreatic lipase | |||
WRYKVSVTLSGKKVTGHILVSLFGNKGNSKQYEIFKGTLKPDSTHSNEFDSDVDVGDLQMVKFIWYNNVINPTLPRVGASKIIVETNVGKQFNFCSPETVREEVLLTLTPC | |||
>PNLIPRP1_homSap LIP-PLAT chr10:118340480 pancreatic lipase-related 1 | |||
WRYGVSITLSGRTATGQIKVALFGNKGNTHQYSIFRGILKPGSTHSYEFDAKLDVGTIEKVKFLWNNNVINPTLPKVGATKITVQKGEEKTVYNFCSEDTVREDTLLTLTPC | |||
>PNLIPRP2_homSap LIP-PLAT chr10:118370455 pancreatic lipase-related 2 | |||
WRYKVSVTLSGKEKVNGYIRIALYGSNENSKQYEIFKGSLKPDASHTCAIDVDFNVGKIQKVKFLWNKRGINLSEPKLGASQITVQSGEDGTEYNFCSSDTVEENVLQSLYPC | |||
>PNLIPRP3_homSap LIP-PLAT chr10:118177414 pancreatic lipase-related protein 3 chr16:70520943 | |||
WRHKLSVKLSGSEVTQGTVFLRVGGAVRKTGEFAIVSGKLEPGMTYTKLIDADVNVGNITSVQFIWKKHLFEDSQNKLGAEMVINTSGKYGYKSTFCSQDIMGPNILQNLKPC</font> | |||
>PLAT_cloPer Clostridium perfringens phospholipase toxin DQ183948 | |||
KELVAYISTSGEKDAGTDDYMYFGIKTKDGKTQEWEMDNPGNDFMTGSKDTYTFKLKDENLKIDDIQNMWIRKRKYTAFPDAYKPENIKVIANGKVVVDKDINEWISGNSTYNIK | |||
>ALOX_8R_pleHom Plexaura homomalla (coral) 8R-Lipoxygenase LOX-8R beta calcium membrane | |||
AIYNVEVETGDREHAGTDATITIRITGAKGRTDYLKLDKWFHNDFEAGSKEQYTVQGFDVGDIQLIELHSDGGGYWSGDPDWFVNRVIIISSTQDRVYSFPCFRWVIKDMVLFPGEA | |||
<font color="purple">>LOXHD1_galGal_PLAT:01 | |||
KVYEVVTVTGDVRGAGTDANVFVTLFGEFGITPKTHLTSKSSTAFERSKTDVFRVKTNNVGQIKKIRIEHDNTGLNAGWFLDRVIVTDMNRPhLRFYFPCNNWLSKEDGDGLYVRDLI | |||
>LOXHD1_galGal_PLAT:02 | |||
NKYVVRVFTGEVSGSGTDADVFINIFGEKGDTGVRKLDNDKDNFEKGAEDKFTLDAPNLGRLRKINIGHNNKGGSAGWFLAKVIIEDIGNKCVYQFPVGRWFALDEDDGKIQRDIL | |||
>LOXHD1_galGal_PLAT:03 | |||
IVYNVAVVTGDIRGAGTNSKIHVILHGSKglKNSGTIFLEGGEFERARTDLFNVEIASLLSPLSRVTIGHDNCGVSSGWYCEKVVVYCPFTGIEQTFPCGKWLDEDEGDGLIERELY | |||
>LOXHD1_galGal_PLAT:04 | |||
NPWSLWIWTSDIKNAGTDATIFFQIYGDKGKSDEMKLDNNSDNFEAGQTDKFMIELPDLGTFYKLRIWHEKRNPFAGWHLDKVTLLKTLTKDKYSFNCGRWLDINEDDNEIVRELP | |||
>LOXHD1_galGal_PLAT:05 | |||
IKYRVTVCTGMVSGSGTDANVFVCLIGDQGDTGDRVLYKCinNVNKFEKGNADEFFVEAVTLKQVRRVRIGHDGKGGSSGWYLAKVIVREEGQPesEAVEFPCYRWLDKNEDDGQIVRELV | |||
>LOXHD1_galGal_PLAT:06 | |||
VSYHISVKTGDIPGASSDSKVFIKLYGEKADTSKETLLVSdndLGNYFERGRVDEFTIDTMDIGKINRILIGHDNVGLRSGWFLASVQITVPVQGRQYMFPCNRWLDKDEADGRVEVEVY | |||
>LOXHD1_galGal_PLAT:07 | |||
INYEVSVVTGDVRAAGTNAKVFMQIYGETGKTELIILENRSNNFERGATDIFEVEAADVGKIYKIRIGHDGKGIGDGWFLESVTLKRLATKmdetdkkkkkkkkkseeeeeeeetkveevmDVYTFVAHRWLAKDEGDKELVVELV | |||
>LOXHD1_galGal_PLAT:08 | |||
NTYEVHVLTGSVWGSGTDANVFLSIYGIErGDTGERQLKRSnNLNKFEKGQVDVFTIKAIDLGELKKLRIRHDNSGSSPSWFLERVEIVDLKESTTYYFPCQRWLAVEEDDGQIVRELV | |||
>LOXHD1_galGal_PLAT:09 | |||
TTYIVKVKTGDKKNAGTDANVFITLYGSKDDTGIVSLKASklNKNKFERGKIDEFTVESVDIGDLKKIKIGHDNAGNSNGWFLEWVEIDAPSLGQCLKFPCGRWLDKSEDDGAIERFIF | |||
>LOXHD1_galGal_PLAT:10 | |||
VPYEITVYTSDIFGAGTDADVFIVLYGSDgicTQQKSLCLNKReQRMYFERNSVNQFIVELEDVGDIIEKIRIGHNGGGLNSGWHLDRVAIRRLLPNgkgsETITFPCERWLAKSEDDGEIIRELV | |||
>LOXHD1_galGal_PLAT:12 | |||
IPYHITVTTGTEYDSSTDSRVFIIIMGPQkVRTERLWLDLPeGKDEFADGSVEKFSVWGLDVGEIKKVEVGHDGATPESCWLMEELTIVVPTKGVMYNFVCKCWLARDKGDGLTSRILN | |||
>LOXHD1_galGal_PLAT:13 | |||
ILYEVTVVTGDIESGGTDAGIFMTVFGSNGNTEEMQLDKNGDRFERGQEDSFIMEIADIAPLRKMRIRTDAKGTRPDWFLERIVMRNLTNQEVATFTYGDWLSKVKNAKGSLVCEM | |||
>LOXHD1_galGal_PLAT:14 | |||
TTYTIQVKTSDIGGAGTDANVSLILFGENGDSGTLALKESnKSNKFERNQMDEFNFPNMLSLGDLCKVRIWHDNKAY | |||
> | >LOXHD1_galGal_PLAT:15 | ||
KAYEIVTVTSNREDAETKENIWIILEGKLGRSKEFLMENSskKRRFERRGSTDTFQFSSKNLGDIAAICVGHCPKDgkkssakADVYWHVKEIIITEMELCNKYFFRCNGKIPLRYKRRDYKVFEC | |||
> | >LOXHD1_galGal_PLAT:16 | ||
VKYETIVVTGFEKGAGTDANVFITIFGLNGDSGKRALKQKFRNLFERGKTNRFYLETLD | |||
</ | MGELKKVRIEHDNSGLAPGWLVERVEITNSATGVTTIFPCGKWLDENRGDGLTWRELF</font> | ||
<span style="color: #0066CC;"> | <span style="color: #0066CC;"> | ||
>LOXHD1_schMan_PLAT:01 Schistosoma mansoni (flatworm) Bilatera; Platyhelminthes; Trematoda XP_002576380 from contig NS_000200 | >LOXHD1_schMan_PLAT:01 Schistosoma mansoni (flatworm) Bilatera; Platyhelminthes; Trematoda XP_002576380 from contig NS_000200 | ||
| Line 468: | Line 654: | ||
>LOXHD1_schMan_PLAT:14 | >LOXHD1_schMan_PLAT:14 | ||
TCLDIFVKTGNMPASSTDANVYVQLFGEYGDSGEILLKQTVSNQKPFQNNSIDHFKIPSILKLGNLARCRIWHDNKGSSPNWYCEWLEVKEVLIPGEKNLACNWKFAFNKWLSVSDDNKQLLRDAPCS | |||
>LOXHD1_schMan_PLAT:15 | >LOXHD1_schMan_PLAT:15 | ||
| Line 474: | Line 660: | ||
>LOXHD1_schMan_PLAT:16 | >LOXHD1_schMan_PLAT:16 | ||
VMYKVSIYTGTKGCANTDANIFITMFSTTPGLNSGRIALKRENNNLFDRKQLDEFYVESIDLESIQRIIIEHDNTGVSPDWYLDKVLITNQTNNQIHLFQCYQWISKKKGDCRLWKELLVS | VMYKVSIYTGTKGCANTDANIFITMFSTTPGLNSGRIALKRENNNLFDRKQLDEFYVESIDLESIQRIIIEHDNTGVSPDWYLDKVLITNQTNNQIHLFQCYQWISKKKGDCRLWKELLVS</span> | ||
</span> | <font color="red"> | ||
>LOXHD1_triAdh_PLAT:01 | |||
VAYQFGVKTGDKKGSDTDAVYIQVIGTKDKIPKKRLFKKQETEKTERGNLFKFDKSTVEKFAVQHRDIGDPVKLIVEHDGNEKRHGWFLEEITLTNIQSKKSWLFPCHKWLSKYEGDRKLCYE LKPLAKAGK | |||
>LOXHD1_triAdh_PLAT:02 | |||
AVYEVSVLTGDKRGAGTDANVSVTLFGKHTSSPKIQLLKSSKHKNPFERNNTDEFKIRTRDVGKLSKIRIEHDNAGFGPGWFLDKVIICNLEKPNVKYYCPCNQWLAKDVGDKSISRD LTAYTDPNAAPSA | |||
>LOXHD1_triAdh_PLAT:03 | |||
YVYIVHTFTGNKRGAGTDANVYAVIFGDSGDTGEKRLDNSKNNFEKSRKDTFKLSCSCVGKLERLRIRHDNTGLFAGWYLDKVVVEDPQEQQSYTFYCRRWLSKTEDDGEICRDLI VSASGDDGDDSVAPKG | |||
>LOXHD1_triAdh_PLAT:04 | |||
YPYHIHVTTSDVKNAGTDAEVYVVMHGEGKKSKELNSGKLVLANSEKKKNTFERAMTDIFHMECAEMLSPLTKLTVGHDNKGLAAGWHLDRaIVIDCPTTGIEQTFLCQQWLDRKAGDGLTERELVEA FDMRKTRRPK | |||
>LOXHD1_triAdh_PLAT:05 | |||
QLWFAWIWTSDIRGAGTDANVSMQIYGDKGKSQEIKLGNNTDNFEQATLDKFKLEIDQVGVPYKLRIGHDNSNAFPGWHLDKVKLENMNDKEQYLFNCNRWLSRSEEDNEIIRELPAS GPNCPNYPI | |||
>LOXHD1_triAdh_PLAT:06 | |||
VIYEVSVHTGNKMGGGTDANVFIKIYGELGDSGYRPLKSSKSHNNKFERNQVDVFHIEAVTLKALKKIKIGHDGNNPGAGWFLDKVVIKELNGEASNEFPCNRWLSKSEDDGQIVRELFLK SDTPLLKT | |||
>LOXHD1_triAdh_PLAT:07 | |||
TSYHISVKTGDVRNAGTDANVFIQIFGAKDDTGRVRLKQSLNTSNKFERNRIDKFIIEAAQIGKIEKIIIGHDGKGLGSGWFLDYIELDVPSVGRLYRFSCHQWFDSTEGDRKVERELYPS ECIKSAAK | |||
>LOXHD1_triAdh_PLAT:08 | |||
IPYQISVHTGDIRHAGTDSNVFAVIYGENGKTEELKLRNKSDNFERGQVDVFKVECEDVGKLRKLRIGHDSAGMGSAWFLDKVYVRRLPPKSGKKSKETDEREEETAKKDADEPEKLDENNYLFVANRWLSKEEGDRQTVIEIS PVGVDGALAE | |||
>LOXHD1_triAdh_PLAT:09 | |||
MTYTIRVITGNKFGCGTNANVFINMYGEEGDSGERQLKKSETHTDKFERNQEDVFKISCLSLGELKKIKIRHDNSGFRPAWFLDKVIIEVGESKYQFMCDRWLAKDEDDGQISRELLPQ SDEQSRAEAIGASKDLQKKVAS | |||
>LOXHD1_triAdh_PLAT:10 | |||
TTYNVSVTTGDIKGAGTDANVHIVLYGEKDDTGLIHLKNSTTHSNKFERNQEDRFVVEAIDIGELKKIKIGHDNKGGMAGWFLNKVEIDIPSLGRRLLFPCGRWIDKGKDDGALER ELYPLNEAEETYRPH | |||
>LOXHD1_triAdh_PLAT:11 | |||
IPYEVTVYTTDKRGASTGANVYVVIYGEENQTEQASLEPDKKRRKQYFKNGAIDKFVLELDDVGEEITKLRIGHDGKGWGAGWHLDKVEICRLLDGGKASKKFTFQCNRWLASDEDDGAIVRELVPS EIVEKSSKDGGQVKTKVTKPTDGLKV | |||
>LOXHD1_triAdh_PLAT:12 | |||
KPYTIHVFTGDVDGAGTNANVFLTIFGESGDSGERKLAKSDTHYDKFERNQEDIFHIEAADLGRLFKVKIRHDNTGSLFSPAWFLNRIEIVDDENEETTAFPCERWLAKKKDDGKIDRTLFVK | |||
>LOXHD1_triAdh_PLAT:13 | |||
VPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVSHNSYMPDSGIYLKEIEVDVPTIGNKYIFPCNRWLAKDKDDSKTSRIFTAA | |||
>LOXHD1_triAdh_PLAT:14 | |||
KPYELTIHTGDVQNAGTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIPLELDDVGTIKKIRIGHDGKGSRTDWYLEKASIQRMDTLDMYMFRANQWFSKKIDDKKLVREI PAETSKEGATTIKK | |||
>LOXHD1_triAdh_PLAT:15 | |||
INYVLSTHTSDKRGSGTDANVFVIIFGENGDSGEIALKKSETNWNKFEKGQTDVFLINDRLSLGRLQKLRIWHDNAGFGASWHLASVDIVDESTGVKYTFPCDKWLSKSNGDKLILREL PCAETAGNTAASKATKQESKSGK | |||
>LOXHD1_triAdh_PLAT:16 | |||
AEYEIAFTTGTEKRAGTNQDVAIVLKGKSEKSREFLIENNEDKKYFSKGKTNKFTYTCKPLGDITKAIVSHRESAIGEEPDSKNSSWYLKVVTVVHKASGTTYKFPCNKWIDLDDDEENNSSVTLKC KSADVAASSKAKPVELKP | |||
>LOXHD1_triAdh_PLAT:17 | |||
VKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* </font> | |||
<span style="color: #00CC66;"> | |||
>LOXHD1_monBre_PLAT:01 Monosiga brevicollis (monosiga) | |||
GHASLEIFTSKRSNPVSKSEKYITVVGTKGRSDAIQLAAPGVHFRAGNKDVFQVNLTGIGKPTKVILENTGTKRTDGWCVSKVVLVKKTDKGTRRYRFSGPVWLSKHHDEMKLKR | |||
>LOXHD1_monBre_PLAT:02 Monosiga brevicollis (monosiga) | |||
NGYRIDCYTGDVANAGTDAVATIQLFGSKGQSPMVELRRSDGQAFQRARVATFTLDDLANLGKLKKLVISHNGHGMASGWFLDKIIVTSLSSNKATVFPCDAWLDRKNGRSKELVAR | |||
>LOXHD1_monBre_PLAT:03 Monosiga brevicollis (monosiga) | |||
TTFTIRVMTGDRRGAGTDANVQCTLFGEDGESGPHTLNTSRNDFRRGHTDVFAVSSRKIGTLKRLRIWHDNGGAGPAWFLDAVEVVDEASGQTYRFECNRWLAKDEDDGQISRELTCN | |||
>LOXHD1_monBre_PLAT:04 Monosiga brevicollis (monosiga) | |||
KSYKLTIFTGDKRNAGTSANVFCKLVGERGASDNVILENSSKNFQRDRTDIFTVEASDLGSLRHIVLGHDNHGMGAGWYVERFSLEVPSEGKLYNVDVKQWFATDMSDGAIERTFRLD | |||
>LOXHD1_monBre_PLAT:05 Monosiga brevicollis (monosiga) | |||
LDWKCTIYTSDVANAGTDANVFMQVYGKKGKTDVVPLKNKSDTFERGQTDELRVQLINVGSLRKLRVWHDNKGMASGWHLDRIVLSRDGEEYIFPCAEWLAVSEGDKEIVRELPAT | |||
>LOXHD1_monBre_PLAT:06 Monosiga brevicollis (monosiga) | |||
VEYTVRVATGHARFAGTNADVFVMLTGELGDSGKRALLRSQTNRNKFERGKEDVFTVAAVDLGKLTSVTVGHNNAGTSAGWFLDKIVVLDPRRGEEEEFPCHRWLAVDADDGQIERELVPK | |||
>LOXHD1_monBre_PLAT:07 Monosiga brevicollis (monosiga) | |||
TTYIVKIKTGDVRHAGTDANVFVQLFGKTGESTQLKLRNSETYSDAFERNKMDIFKFELLDLGDLSRILVGHDNKGMGAAWFLDYVEVEVPSIRTRWKFPCSRWFSKSQDDGLTER | |||
>LOXHD1_monBre_PLAT:08 Monosiga brevicollis (monosiga) | |||
APYLFRFYTSDVAFAGTDANVSVVLYGDEGKTEELVVNNQSDNFERGKADDFKLACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQARIYTFEHNDWLRKGTKAKPFMVEL | |||
>LOXHD1_monBre_PLAT:09 Monosiga brevicollis (monosiga) | |||
RTYRVKVHTGDQKGAGTDANVYVNLHGSLGDSGDRHLKNSLTHTNKFQRKTVDEFDIDAVTLGDINKVKVWHDNAGLGAAWYLEKIEVVDTADDKTYIFPCAQWFAKSMGDGQIAREL | |||
>LOXHD1_monBre_PLAT:10 Monosiga brevicollis (monosiga) | |||
FKYRISVHTSDVKHAGTDANVDIVLYGEKGDTGKIRLAKSETHRDMWERGNCDVFTVSAIELGDLKRVDIMHDGKGVGSGWHLNKVVVDAPQAGKTWTFMCDAWLDKATDDGTMAK | |||
>LOXHD1_monBre_PLAT:11 Monosiga brevicollis (monosiga) | |||
VPYEIIIKTSDVRNAGTDANVFIDLYGRDQEERDLTAHHEFKDAVKAHFERNLEDRFNVELPDVGSIYKIRLGHDGKGMSSSWHVASVVVINQRTHERFEFPCDAWLSKDKDDKKLVREFAVG | |||
>LOXHD1_monBre_PLAT:12 Monosiga brevicollis (monosiga) | |||
AIYKIHVFTGDIKHAGTDANIYVQIFGDTGDSGEIKLEKSETYRDKFERGHEDIFTHRCLDLGPLRKIKVRSDGKGLMGGDWYLDRVEVHQENDLSEPPVRFVCQDWFKRGKQEGDTLEREITAQ | |||
>LOXHD1_monBre_PLAT:13 Monosiga brevicollis (monosiga) | |||
LEYKVTVYTGTDTQAGTTANVWLQLFGEKDAPLTPAPPSPSKLSRSGSLFGRRRRASSDAQSVSSSLSAGASGPRETSTGRLQLNNAPADLQSGAKTTFTVTGLDVGELVGLEIGHDDERDKWYLEQVVVEVPKSATHAARRYEFKAGVWLAARADGSTSGSAK | |||
>LOXHD1_monBre_PLAT:14 Monosiga brevicollis (monosiga) | |||
LEYTLKVYTASADGAGCTAVPQVQLFGDKHTTEALPLRAGGDILPASVVETQHRVPDLGALLKVRLMVPRGSSWTVEKVEFGRAGQTPITFVGSDGSAVTLGAERLS | |||
>LOXHD1_monBre_PLAT:15 Monosiga brevicollis (monosiga) | |||
TSYRVYVTTADERGTGTDANVSIILYGAMGDSGEHSLTKSETFDDPFERGNTDVFTLEVPDLGELQRARIWHDGKGMFSSWKLDKIVVVVEATQSRYELPCGQWLSKNKGDKQLTRDLAVASKR | |||
>LOXHD1_monBre_PLAT:16 Monosiga brevicollis (monosiga) | |||
GTYKLEVATDSRAGGGCKGPVRIMLLDAEKNQLPLTLEPPGGEFAPGSVEHLVFDNVMLLGPLTELRIRRKPSGASSRRGAEDDDEDDNEASGASSQSGSAVSPWHLEHIIVKHLQSGQSFVFKGPSKGLSRSRAKLSVHTEA | |||
>LOXHD1_monBre_PLAT:17 Monosiga brevicollis (monosiga) | |||
AQYEVAVTTGTERGAGTDSNVFVTLFGKNGDSGERALAKSKTFRNMFESGNTDVFDVECQDLGELTKIEVKSDLKGFGAAWQLDKIKVTRTGSQNSWQFKCDQWFDKKQGAEHTFSVA</span> | |||
=== Reference PLAT:07 domains === | === Reference PLAT:07 domains === | ||
>PLAT:07_homSap Homo sapiens (human) | >PLAT:07_homSap Homo sapiens (human) | ||
2 LVHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQ 0 | 2 LVHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQ 0 | ||
| Line 563: | Line 820: | ||
0 LACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQARIYTFEHNDWLRKGTKAKPFMVELPLRRIETVDDNGREVVEELALDANKRTYR 1 | 0 LACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQARIYTFEHNDWLRKGTKAKPFMVELPLRRIETVDDNGREVVEELALDANKRTYR 1 | ||
'''See also:''' [[USH2A_SNPs|Usher: USH2A]] | [[CDH23_SNPs|Usher: CDH23]] | [[Opsin_evolution:_RBP3_%28IRBP%29|RBP3 (IRBP)]] | [[Opsin_evolution:_RPE65|RPE65]] | [[Opsin_evolution:_transducins|Transducins]] | |||
[[Category:Comparative Genomics]] | [[Category:Comparative Genomics]] | ||
Latest revision as of 16:44, 27 December 2009
See also: Usher: USH2A | Usher: CDH23 | RBP3 (IRBP) | RPE65 | Transducins
Introduction


LOXHD1, a large mis-annotated autosomal gene consisting solely of 16 concatenated 120-residue PLAT domains (named for polycystin/lipoxygenase/alpha-toxins), recently surfaced as a causative gene for age-progressive postlingual non-syndromic hearing loss type DFNB77. Of nearly a hundred known deafness genes, only MYO3 (myosin IIIA) and DFNB59 (protein product pejvakin -- echo in Persian) share this autosomal recessive progressive phenotype.
Howver there is no evidence to suggest that the three encoded proteins physically interact within auditory hair cells nor that heterozygotic combinations give rise to digenic disease (as argued in Usher Syndrome). Further, with larger numbers of pedigrees, each might exhibit syndromic effects (eg affect vision or kidney function).
Mouse knockouts have not been studied; delicate auditory hair cells may just be exquisitely sensitive detectors of subtle disruptions of gene function. While LOXDH1 is specific to hair cell stereocilia within the cochlea, GenBank transcripts also imply expression somewhere within testis, placenta, hippocampus, cerebellum cortex, medulla, stomach, liver, lung, and bone marrow. The protein has not been ultrastructurally localized in any of these other tissues.

Some of these transcripts are exon-skipping, for example AK302484 disables the 12th PLAT domain though retains downstream reading frame. It's unclear whether such transcripts are functional or just splicing error inevitable in a giant protein (41 exons, 2273 amino acids) and merely destined for immediate turnover. The skipped exon, LGHDGASPESCWLVEELCLAVPTQGTKYML NCNCWLAKDRGDGITSRVFDLLDAMVVNIGVK, is still 72% conserved from human to shark; nothing other than functionality could explain such conservation.
Mutations do not surface during cochlear development but rather in mature stereocilia, which degenerate over time. LOXDH1 binds to the inner plasma membrane of stereocilia, perhaps organizing protein binding there or stabilizing some aspect of the membrane. As with other concatenated domain proteins (see CDC23 and USH2A, subtle dysfunction in one of many internally repeated domains surprisingly cannot be compensated for by the remaining intact domains.
Here the mouse I1342N disrupts the hydrophobic pocket between two beta sheets of the tenth PLAT domain but the protein is still made and localized properly. The first morphological change is observed at mouse day 21: fused stereocilia and membrane ruffling at the apical hair cell surface in the basal cochlear turn. Secondary effects arise later and include damage to spiral ganglion neurons.
Human allele R670* would eliminate all but the first five PLAT domains (of 16 total), possibly producing a normal but shortened protein, but might not produce protein at all due to nonsense-mediated decay. These individuals do not experience tinnitus, balance problems or vertigo, suggesting normal vestibular function even though LOXHD1 is expressed there too.
One difference between cochlear and vestibular hair cells is retention of the apical kinocilium in the latter. This is lost in developing cochlear hair cells as stereocilia mature. This correlates with peak LOXHD1 expression, suggesting to Grillet et al that LOXDH1 might help stabilize the stereociliary bundle in the absence of kinocilium.
Recall the kinocilium is a non-motile cilium-like structure present in the crista ampullaris of the semicircular ducts and the sensory maculae of the utricle and saccule but not in the cochlear duct organ of Corti (where in mammals it degenerates following maturation of the stereociliary bundle). Since retinal receptor cells are modified cilia and LOXDH1 functions most critically in microvillar hair cells after the kinocilium is gone, a syndromic effect extending to vision would not arise based on kinocilium/photoreceptor cilia homologization.
Phylogenetic occurence
The PLAT domain is quite ancient, readily tracing back to prokaryotes. However in other proteins it occurs only as single copy, often in conjunction with one or more catalytic domains. A common denominator in PLAT domain proteins is their association with lipid or membrane metabolism. This is perhaps mediated by extruded tryptophans which extend out from the standard beta barrel surface.
LOXHD1 itself has highly conserved orthologs throughout vertebrates, indeed deuterostomes (though the Branchiostoma gene model XM_002606291 contains other domains). These species all contain mechanosensory cells with ciliary F-actin based protrusions though expression has not be localized outside of mammals. No counterparts of the gene can be found in Ecdysozoa (strictly non-cilary mechanosensation) yet lophotrochozoa (Schistosoma and likely Aplysia) have full length counterparts, as can cnidrians (Hydra, Nematostella) though gene models at this time are fragmentary.
This establishes gene loss in the ancestral ecdysozoan, rather than gene innovation in Bilatera. However the overall impression is phylogenetically spotty occurence outside of deuterostomes. It's unclear whether some other gene product can compensate for lost LOXHD1 or whether it simply does not play an essential role.
Most surprisingly, full length orthologs of LOXHD1 with 17 PLAT domains and very similar intronation patterns are readily recoverable from early eukaryote genomes, notably Trichoplax and the choanflagellate Monosiga. This seems to rule out an obligatory association with F-actin-based mechanosensory organelles as the basal metazoan Trichoplax exhibits only 4 distinct cell types and the early animal Monosiga is unicellular.
Thus LOXDH1 is not even a metazoan innovation. Lateral gene transfer makes no sense in this context and is contradicted by the gene tree. The extraordinary conservation Monosiga to human -- surely in the 95th percentile of all genes -- establishes a long-selected intracellular role.
In the plant world, occurence is restricted to Chlamydomonas and Micromonas proteins (XM_001703369, XM_002502455) with 20 PLAT domains and numerous WD40 domains (ie not necessarily orthologous to animal pure poly-PLAT proteins). A curious note on the Chlamydomonas genbank entry states "identified by comparative genomics as being present only in organisms having motile cilia." Vertebrate kinocillia are non-motile.
Within deuterostomes, the large length of the gene does not sit well with gappy, incomplete and sometimes chaotic assemblies. Chicken, lamprey, amphioxus, tunicate and sea urchin are among the many that do not yield satisfactory full length gene models, in part because transcript support is limited to small regions of large proteins. However these species do contain long poly-PLAT proteins orthologous to human LOXHD1 (based on consecutive domains having consecutive best-blast).
Some of these unreliable sequences are provided in the reference sequence compilation below. Ironically, the obscure Monosiga and Trichoplax have better assemblies and more accurate gene models presumably because their genomes are much smaller.
Thus within vertebrates it is only feasible to compare selected PLAT motifs. Here there is a risk of using best-blast because significant cross-over matches will occur to other PLAT domains within the same protein, so some additional signature must be used for validation.
PLAT domain structure
The structure of three proteins containing single PLAT domains have been determined. The best blastp of human LOXHD1 to these is a coral lipoxygenase/allene oxidase natural fusion protein with PDB accession 3DY5. Lower quality but still useful matches occur with RAB6 (accession 3CWZ) and rabbit reticulocyte lipoxygenase (accession 2P0MA).
The basic structure of a PLAT domain consists of a sandwich of two 2-stranded beta sheets (beta barrel). The lipoxygenases are calcium-binding via extended pairs of acid residues and have conserved hydrophobic residues likely inserting into the plasma membrane but neither these nor the hydrophobic residues are conserved in LOXHD1.
The independently folding PLAT domain is sometimes denoted as LH2 or 'C2-like' depending on the structural publication and domain tool. This introduces confusion because SMART/Pfam/InterPro utilize very distinct HMM models for C2 and PLAT/LH2.
The secondary structure here is so generic that convergent evolution (non-homologous origins) must be considered, as with TIM proteins. Here however significant blastp matches at the primary sequence level unify PLAT and LH2 sequences to the exclusion of the entire C2 family. This agreement cannot have originated repeatedly by chance given the vast combinatorial space available to amino acid sequences.
Further, all PLAT/LH2 domains share characteristic conserved residues. Thus PLAT and LH2 domains are in fact all homologous (descended from a single source) and so multiplicity of domain names is inappropriate. PLAT is used here rather than LH2 (lipoxygenase homology) because the former is a more comprehensive acronym of protein classes containing the domain (lacking only lipases).
C2 domains bear an uncertain relationship to PLAT domains because a resemblance occurs only at the secondary and tertiary structural levels. The lack of convincing blastp of HMM profile matches could be due either to extreme divergence of primary sequence or to convergent evolution resulting in the same basic fold (and perhaps a similarly convergent hydrophobic intermediating function). If ever homologous, PLAT and C2 diverged in prokaryotic times.
The name C2 implies calcium binding yet even here some C2 domains function independently of calcium. It isnot clear whether calcium binding is ancestral or derived. If ancestral, it has not been resolved whether binding has been lost once or multiple times in different regions of the domain tree. While PLAT domains can be structurally aligned to C2 domains, that is potentially misleading and unnecessary given three determined structures for bona fide PLAT domains.
LOXHD1, which tracks back to the earliest unicellular animals, lacks capacity for calcium binding in all 16 of its domains in all descendant clades based on non-conservation of acidic liganding residues relative to coral lipoxygenase, human LOX5 and clostridial alpha-toxin PLAT domains.

PLAT domain evolutionary history

The human genome contains 20 other proteins with significant blastp matching to the PLAT domain of LOXHD1. These are shown below along with their flanking domains (GPS, RUN etc). After adjusting for gene expansions (paralogy), only 4 classes occur: polycystins, lipoxygenases, lipases, and RAB6-interacting. These all contain a single PLAT domain. The final column shows its best-blastp to LOXHD1; no strong pattern emerges though perhaps PLAT:09 may conserve the most ancestral features and PLAT:15 the least.
The first two columns at bottom shows the blastp score of a given human LOXHD1 PLAT domain averaged over it match to the 12 other genes; the second two columns show the average match of other genes over the 16 LOXHD1 PLAT domains.
The domain tree thus looks like ((((LOXHD1,RAB6Is),polycystins),lipoxygenases),lipases). A possible parsimonious evolutionary scenario postulates a single standalone PLAT domain originally serving as an auxillary protein to various enzymes with hydrophobic substrates. Later, copies fused with these catalytic domains in separate events with gain in efficiency.

Observe the PLAT domain is N-terminal to the catalytic domain of lipoxygenases but C-terminal to it in lipases and sandwiched internally by a repeated domain in RAB6I, indicating three separate fusion events. These fusion proteins then expanded copy number to the small paralogous gene families seen today. The individual PLAT domains then evolved at different rates, though they continued to share some common constraints (beta barrel, membrane fingers).
The standalone gene eventually expanded to a poly-PLAT protein; today the standalone domain itself is gone (no longer represented in extant species). It is not easy to discern intermediate events: each species seems to generate a unique internal domain tree (above). In other words, individual PLAT domains may have evolved at different rates in different clades, obscuring initial relationships from order of duplication.
Intermediate events may have included 1 domain tandemly duplicating to 2 and then to 4, 8, and finally 16 but other scenarios involving inhomogeneous recombination (and even higher multiplicities) are equally plausible. However the number of repeats has been very stable over many billions of years of branch length; each individual repeat seems important too in view of known point mutations.

PKD1L1 GPS-TM-PLAT-TM chr07:47780815 polycystin 1-like 1 pdb:3CWZ PLAT:14 36%
PKD1 GPS-TM-PLAT-TM chr16: 2078712 polycystin 1 pdb:3CWZ PLAT:01 38%
PKD1L2 GPS-TM-PLAT-TM chr16:79691985 polycystin 1-like 2 signal pdb:3CWZ PLAT:11 35%
PKD1L3 GPS-TM-PLAT-TM chr16:70556160 polycystic kidney disease pdb:3CWZ PLAT:01 40%
PKDREJ GPS-TM-PLAT-TM chr22:45030224 receptor egg jelly signal pdb:3CWZ PLAT:11 32%
DENND5A RUN-PLAT-RUN chr11: 9116949 RAB6 interacting 1 pdb:3CWZ PLAT:09 42%
DENND5B RUN-PLAT-RUN chr12:31426424 RAB6 interacting 2 pdb:3CWZ PLAT:09 37%
ALOX5 NH2-PLAT-lipox chr10:45189635 arachidonate 5-lipoxygenase pdb:2P0M PLAT:01 37%
ALOX15 PLAT-lipox chr17: 4480963 arachidonate 15-lipoxygenase pdb:2P0M PLAT:03 26%
ALOX12 PLAT-lipox chr17: 6840108 arachidonate 12-lipoxygenase pdb:2P0M PLAT:09 30%
ALOX15B PLAT-lipox chr17: 7888129 arachidonate 15-lipoxygenase pdb:2P0M PLAT:02 34%
ALOX12B PLAT-lipox chr17: 7916679 arachidonate 12R-lipoxygenase pdb:2P0M PLAT:09 29%
ALOXE3 PLAT-lipox chr17: 7939943 arachidonate lipoxygenase 3 pdb:2P0M PLAT:16 32%
LIPC LIP-PLAT chr15:56511467 lipase C pdb:2P0M PLAT:11 30%
LIPG LIP-PLAT chr18:45342425 endothelial lipase pdb:2P0M PLAT:09 32%
LPL LIP-PLAT chr08:19840862 lipoprotein lipase pdb:2P0M PLAT:11 24%
PNLIP LIP-PLAT chr10:118295418 pancreatic lipase pdb:2P0M PLAT:16 24%
PNLIPRP1 LIP-PLAT chr10:118340480 pancreatic lipase-related 1 pdb:2P0M PLAT:16 27%
PNLIPRP2 LIP-PLAT chr10:118370455 pancreatic lipase-related 2 pdb:2P0M PLAT:01 31%
PNLIPRP3 LIP-PLAT chr10:118177414 pancreatic lipase-related 3 pdb:2P0M PLAT:02 27%

Below all PLAT domains found in human proteins are aligned with the 16 domains of human LOXHD1. The coral lipoxygenase is included because it provides the template for beta strand assignment and locating the problematic calcium binding and hydrophobic extended residues. The lower alignment is just with human LOXHD1 domains and uses an optimized seventh PLAT domain (the KE expansion is removed).

Individual PLAT domains can be quite conserved over the mammalian time scale. That is illustrated by a difference alignment of the fifth PLAT domain in 40 vertebrates (human to lamprey) below. Note columns where human sequence represents a unique change -- such as H->R in column 5 -- may be minor alleles or disease variants rather than the predominate residue in the overall human population.
01.hg18_6 MARYHVTVCTGELEGAGTDANVYLCLFGDVGDTGERLLYNCRNNTDLFEKGNADEFTIESVTMRNVRRVRIRHDGKGSGSGWYLDRVLVREEGQPESDNVEFPCLRWLDKDKDDGQLVRELLPSDSSATLK 02.panTro ....R.............................................................................................................................. 03.gorGor S..R...........................................................................--------------------------..................G...... 04.ponAbe ....R..............T.....................................................................................-....................N.... 05.rheMac ....R.........................................................................................................................N.... 06.calJac ....R...........................................................K.............C...............................................N.... 07.micMur ....R...........................................................M.............................................................N.... 08.otoGar ....R. ............K.................-..........H.P.V.......T....................N.... 09.tupBel ...R...........................................................K............A................................................N.... 10.mm9_6_ ....R...........................................................K.....V.......................................................N.... 11.rn4_6_ ....R...........................................................K.....V........................................Q................... 12.dipOrd ....R.....................Y..................E..................K.....V.......S.S.............................................N.... 13.cavPor ....R.......F.............Y...................................L.K.............................................................N.... 14.speTri .V..R.....................Y......-.....-------------............K............P......E...L.....................................N.... 15.oryCun ..........S.............K..........................................................TG.N.... 16.ochPri ....R...........................................................K.............S....V..........................................N.... 17.vicPac ...R.......V.............................I.....................K.................F.E.........................................N.... 18.turTru ....R...........................................V...............K.....V...........F.......................---------...........N.... 19.bosTau ....R.....................Y.....................................K.................F.E....................-....................N.... 20.equCab ....R...........................................................K............A....F.E.........................................N.... 21.felCat ....R...........................................................K............G....F.E.........................................N.... 22.canFam ....R...........................................................K............G....F.E.........................................N.... 23.myoLuc ...HR......D.R......S.....Y............T.K..DEM.Q...............K........T.V.G....F..................Y........................N.... 24.pteVam ...R.........................................M.................K..........R.G....F.E.........................................N.... 25.eriEur ....R.....................Y.....................................K............G....F.A..............M..........................N.... 26.loxAfr ..Y.......-..M.....................A..L.K............G......E.........................................N.... 27.proCap ....R.....................Y..A.......M...........R.........A..L.K............A......E..............M...---...................NN.... 28.echTel ....R.....................Y..........MF.....V..............A..L.K......M.....A......-...LKK.........K..G.G....................N...R 29.choHof .V..R.....................Y.........W.........M....S...Y...A..L.K.K.I.VG.....GN...F.E..............M....N...AR.Q.....I......G.N.... 30.monDom ..Q.R..L...DI.....N.QAFV.....A......M.......MEICQ..........A..V.K......G.....K....F.VK..................N..F...Q..............NSR.. 31.ornAna IVK.R...V..D.N......R.FI..I.........I.........T.........FV.A..LKQ......G.....GS.....AK.I.........EA.....Y.....NE....I....V.AGE.PL.. 32.galGal VIK.R......MVS.S......FV..I..Q....D.V..K.I..VNK.........F..A..LKQ......G.....GS.....AK.I.........EA.....Y.....NE....I....V.AGE.PL.. 33.anoCar IK.R...H..NVS.S....H.FA..I..Q....D.V.Q.SV.TVNK....S....I..A.NLKQ......G.....GS.....GK.I...D.....EAQ....Y.....NE....II...V.AGR.SL.E 34.xenTro ...F.IE.K..MNK.......D.I..A..LKQ.K....G.....G.C...M.K.V...D.K..TEAI....N..F.RNE...HII...V.AGD.QN.R 35.tetNig LK.RI.I...NVG.G....S.F.NII..L......SMITSK..VNK.....H...L....CLGR.S...VG...R.G.C..F..K.T.........SAI....F....HNE....I....V.AGDGRLF 36.fr2_6_ LIK.R..I...NVS.G....S.F.NVI..L.......MIMSK..VNK.....H...L....SLGQ.....VG...R.G.C..F..K.M.........SAI....F....HNE....I....V.A 37.gasAcu LIK.R..I...NVS.S....S.F.NVI..L.......MFMSK..VNK.....H...L....SLGQ.....VG...R.G.C..F..K.M...D.....LAT....F....RNE....I....V.AGDGRLF 38.oryLat IK.R..I...TVS.S....S.F.NVI..L.......M.MSK..VNK.....H...L....WLGQ.....VG...R.G.C..F..K.M.........AAI....N....QNE....II...V.AGEG 39.danRer VK.R...Y..DVS.S....H.F...I..L....D.S.I..K..INK..R......I..A.SIGP......G...R.G.C.....K.TI.....A..QA.....S..F.RNE....I....Q.FADGRLY 40.petMar VK.L..IH..KHG.G......FV..Y.EQ....D.M..TS...VNK..R..V...V..C..LKR.T....G...R.GS...F..KIM.K-D.RRQCEE.....N....D.E....I....VAGGCQMLK Consensus ...yrvt..tg...g.gtda.v..cl.gd.gdtger.ly.crnn...fekgna...t..s.t$rk.r...!r...k.ggs..y$d.!lvre#.qpes.nv...c.r.l.k#k....l!...l.sgsnatlk
Expansion of the PLAT:07 domain
The seventh PLAT domain (human numbering) is unique domains in having a large compositionally simple expansion in the middle of the domain. It is a unique signature of the LOXHD1 orthology class that would distinguish it from other poly-PLAT domain concatenates of independent origin. That issue is completely hypothetical because no other protein in any vertebrate genome contains more than a single PLAT domain. Indeed, the LOXHD1 gene is single-copy in all species that retain it (including teleost fish). Thus even with very ancient eukaryotic divergencs, the corresponding genes can be taken as straightforward human orthologs.
Presumably a loop out of the beta barrel, the expansion consists of an extremely high AG content at the dna level and encodes charged residues, primarily lysine K and glutamate E residues. These are asymmetrically distributed with the lysines N-terminal and the glutamates distal. The region will be called the KE loop of PLAT:07 here.
Despite minimal net charge, this region may not be able to interact with (hydrophobic) membranes. Yet the split PLAT domain retains the potential to form a standard beta barrel. The seventh PLAT domain is quite variable even within mammals and must be hand-annotated to ensure accuracy (because its quasi-repetitive nature wreaks havoc with alignment). This section uses comparative genomics within eukaryotes to answer the following questions:
- when did the high GA section arise in phylogenetic time? [early eukaryotes]
- what is its source? [internal expansion, replication slippage]
- did it arise from intron or retroposon retention? [species lacking the repeat also have a single exon]
- can the boundaries of 7th PLAT domain be better located? [conservation on both sides of the KE region allow definition of the PLAT domain segments]
- is the 44-species alignment accurate in this region? [corrections are needed when longer than human, eg platypus]
- does the KE domain disrupts PLAT function? [evidently not: the split domain is highly conserved and can form a normal beta barrel]
- is it a mammalian innovation or an unwanted intrusion? [this ancient expansion has variable expansion and contraction in different clades]
homSap LEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLRQLLKKERLKAKLQRKKKKRKGSDEEDEGEEEESSSSEESSSEEEEMEEEEEEEE FGPGMQEVIEQHKFEAHRWLARGKEDNELVVELVPAGKPGPE 1 ornAna LEATDVGEIYKVRLGHSGEGFGSGWFIESLVLKRLVLKEVEPNPEEEKRKAKERERAREQRRKERLKAKQQRKKKKKMKKSSDDEDSEAEDSEEEEGSSEEESSSSSSEEEVEEEFGPGIKEVIDVYKFEAHRWLARDEDDKELIVELEPANRPGPE 1 galGal VEAADVGKIYKIRIGHDGKGIGDGWFLESVTLKRLATKMDETD KKKKKKKKKSEEEEEEEE TKVEEVMDVYTFVAHRWLAKDEGDKELVVELVPDGESELE 1 taeGut REAADVGKIYKIRIGHDGTGIGDGWFLESVTLKRLATKTEGSD KKKKKKKKSEEEETKE EEGMDVYTFVAHRWLAKDEGDKELVVELVPDGESDLE 1 anoCar VEAVDVGKVYKIRIGHDGKGFGDGWFLDSVVVKKLPTKVP KKKKKKKKKKTPEEEEAEE GPGIMEVYNFTPCRWLASDEEDKELVVELVPDEGSELE 1 oryLat IEALDVGKIYKIRIYHDGSGIGDGWFLETVDIKRLTMALVQVEV KKEEAPKKDKKKDKKKKKKEEEEVEIIEE MQEVVETFTFTCNRWLARDEEDGEIVVELLTEENEDLE 1 gasAcu IEAKDVGKIFKIRIGHDGSGIGSGWFLETVDVKRLILALVPKEK KKEDKKKKKKKKEDVDEEGGEE MQEVVLTYSFPCSRWLAGGEEDGELVVELLPDDAKELE 1 takRub IEAKDVGKIFKIRIGHDGLGIGSGWFLEKVYVKHLIMALVPREN KKDDKKKKKKKKKDKEDEEEVGGEE MQEVVVTYHFPCSRWLASGEDDDDLVVELLPEDAEELE 1 petMar VEAMDVGKVVKLRVGHDNSGMGSGWFLDSIVIRRLRQSSPHRPQPVDA EEDEDEEDDEEAEDED VQTYTFPCKRWLARDEDDGEIVRELLPQDCAEME 1 sacKow IEAADVAMLTKIRIGHDNSGRSAGWYLERVIIERFPPKRKM KRKRSGTPRRRGEYDEEDYDD IPETNVVNFVCNRWFAKDEEDHQIVRELLPTDEEALKGH 1 triAdh VECEDVGKLRKLRIGHDSAGMGSAWFLDKVYVRRLPPKSGKKSKET DEREEETAKKDADEPEKLDE NNYLFVANRWLSKEEGDRQTVIEISPVGVDGALA 1 monBre LACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQAR IYTFEHNDWLRKGTKAKPFMVELPLRRIETVDDN 1
Introns relative to PLAT domains
Here it emerges that PLAT domains correspond to 2-3 consecutive exons with conserved phasing, allowing for potential alternative splicing that reduced domain count while not introducing premature stop codons. This may reflect the origin of the 16 PLAT domains by iterated inhomogenous recombination. (to be continued).
Reference sequences (intronated)
The sequences below are broken into individual exons with reading phase (codon overhang) indicated by 012. Alternating colors show odd and even numbered PLAT domains. The 7th domain (which contains the charged KE expansion loop in mammals) is shown in red. In some cases, the PLAT domains are numbered and exon phases are not indicated. In the case of poor quality but phylogenetically important sequences, an approximate sequence is shown.
>LOXHD1_homSap_full 41 exons|2273 aa|chr18|span 179,559 bp|no signal peptide|no conserved cysteines|16 PLAT domains|EIW --> WNW mouse deafness 0 MMPQKKRRRKKDIDFLALYEAELLNYASEDDEGELEHEYYKAR 1 2 VYEVVTATGDVRGAGTDANVFITLFGENGLSPKLQLTSK 2 1 SKSAFEKGNVDVFRVRTNNVGLIYKVR 2 1 IEHDNTGLNASWYLDHVIVTDMKRPHLRYYFNCNNWLSKVEGDRQWCRDLLASFNPMDMPR 1 2 GNKYEVKVYTGDVIGAGTDADVFINIFGEYGDT 1 2 GERRLENEKDNFEKGAEDRFILDAPDLGQLMKINVGHNNKGGSAGWFLSQ 0 0 IVIEDIGNKRKYDFPLNRWLALDEDDGKIQRDILVGGAETT 1 2 AITYIVTVFTGDVRGAGTKSKIYLVMYGARGNKNSGKIFLEGGVFDRGRTDIFHIELAVLLSPLSRVSVGHGNVGVNRGWFCEK 0 0 VVILCPFTGIQQTFPCSNWLDEKKADGLIERQLYEMVSLRKKRLK 1 2 KFPWSLWVWTTDLKKAGTNSPIFIQIYGQKGRTDEILLNPNNKWFKPGIIEKFR 0 0 IELPDLGRFYKIRVWHDKRSSGSGWHLER 0 0 MTLMNTLNKDKYNFNCNRWLDANEDDNEIVREMTAEGPTVRRIMG 1 2 MARYHVTVCTGELEGAGTDANVYLCLFGDVGDTGERLLYNCRNNTDLFEKGN 0 0 ADEFTIESVTMRNVRRVRIRHDGKGSGSGWYLDRVLVREEGQPESDNVEFPCLR 2 1 WLDKDKDDGQLVRELLPSDSSATLK 1 2 NFRYHISLKTGDVSGASTDSRVYIKLYGDKSDTIKQVLLVSDNNLKDYFERGRVDEFTLETLNIGN 0 0 INRLVIGHDSTGMHASWFLGSVQIRVPRQGKQYTFPANRWLDKNQADGRLEVELYPSEVVEIQK 1 2 LVHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQ 0 0 LEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLRQLLKKERLKAKLQRKKKKRKGSDEEDEGEEEESSSSEESSSEEEEMEEEEEEEEFGPGMQEVIEQHKFEAHRWLARGKEDNELVVELVPAGKPGPE 1 2 RNTYEVQVVTGNVPKAGTDANVYLTIYGEEYGDTGERPLKKSDKSNKFEQGQ 0 0 TDTFTIYAIDLGALTKIRIRHDNTGNRAGWFLDRIDITDMNNEIT 2 1 YYFPCQRWLAVEEDDGQLSRELLPVDESYVLPQSEEGRGGGDNNPLDNLALEQK 1 2 DKSTTFSVTIKTGVKKNAGTDANVFITLFGTQDDT 1 2 GMTLLKSSKTNSDKFERDSIEIFTVETLDLGDLWKVRLGHDNT 1 2 GKAPGWFVDWVEVDAPSLGKCMTFPCGRWLAKNEDDGSIIRDLFHAELQTRLYTP 1 2 FVPYEITLYTSDVFAAGTDANIFIIIYGCDAVCTQQKYLCTNKREQKQFFERKSASRFIVE 0 0 LEDVGEIIEKIRIGHNNTGMNPGWHCSHVDIRRLLPDKD 0 0 GAETLTFPCDRWLATSEDDKKTIRELVPYDIFTEKYMKDGSLRQVYKEVEEPLD 1 2 IVLYSVQIFTGNIPGAGTDAKVYITIYGDLGDTGERYLGKSENRTNKFERGT 0 0 ADTFIIEAADLGVIYKIKLRHDNSKWCADWYVEKVEIWNDTNEDEFLFLCGRWLSLKKEDGRLERLFYEK 0 0 EYTGDRSSNCSSPADFWEIALSSKMADVDISTVTGPMADYVQEGP 1 2 IIPYYVSVTTGKHKDAATDSRAFIFLIGEDDERSKRIWLDYPRGKRGFSRGSVEEFYVAGLDVGIIKKIE 0 0 LGHDGASPESCWLVEELCLAVPTQGTKYMLNCNCWLAKDRGDGITSRVFDLLDAMVVNIGVK 0 0 VLYEMTVWTGDVVGGGTDSNIFMTLYGINGSTEEMQLDKKKAR 2 1 FEREQNDTFIMEILDIAPFTKMRIRIDGLGSRPEWFLER 0 0 ILLKNMNTGDLTMFYYGDWLSQRKGKKTLVCEMCAVIDEEEMMEWTSYTVAVKTSDIL 1 2 GAGTDANVFIIIFGENGDSGTLALKQSANWNKFERNNTDTFNFPDMLSLGHLCKLRVWHDNK 1 2 GIFPGWHLSYVDVKDNSRDETFHFQCDCWLSKSEGDGQTVRDFACANNKICDELEET 1 2 TYEIVIETGNGGETRENVWLILEGRKNRSKEFLMENSSRQRAFRK 2 1 GTTDTFEFDSIYLGDIASLCVGHLAREDRFIPKRELAWHVKTITITEMEYGNV 2 1 YFFNCDCLIPLKRKRKYFKVFEVTKTTESFASKVQSLVPVKYEVIVTTGYEPGAGTDANVFVTIFGANGDTGKRELKQKMRNLFERGSTDRFFLETLELGELRKVRLEHDSSGYCSGWLVEKVEVTNTSTGVATIFNCGRWLDKKRGDGLTWRDLFPSV* 0 >Gallus gallus gappy genome,flawed XM_425221 15 PLAT domains MKETKKNKAEEEEEEEEEEEEENEEVVEDVVEEAVEEDGEDESQQKKSKKKSKSEDSEDDGEVKKKKKKKKKTKRKEYSSEEEDDYERRKKKKKKGKKSK EKKKKKGDKSKKGKKEDNFLELYEQELRDYHSDSSNATEDEYNKKKVYEVVTVTGDVRGAGTDANVFVTLFGEFGITPKTHLTSKSSTAFERSKTDVFRV KTNNVGQIKKIRIEHDNTGLNAGWFLDRVIVTDMNRPHLRFYFPCNNWLSKEDGDGLYVRDLIGSLNPMDVPKINKYVVRVFTGEVSGSGTDADVFINIF GEKGDTGVRKLDNDKDNFEKGAEDKFTLDAPNLGRLRKINIGHNNKGGSAGWFLAKVIIEDIGNKCVYQFPVGRWFALDEDDGKIQRDILVGGTEATGIV YNVAVVTGDIRGAGTNSKIHVILHGSKGLKNSGTIFLEGGEFERARTDLFNVEIASLLSPLSRVTIGHDNCGVSSGWYCEKVVVYCPFTGIEQTFPCGKW LDEDEGDGLIERELYEMVSLRQRRLKKNPWSLWIWTSDIKNAGTDATIFFQIYGDKGKSDEMKLDNNSDNFEAGQTDKFMIELPDLGTFYKLRIWHEKRN PFAGWHLDKVTLLKTLTKDKYSFNCGRWLDINEDDNEIVRELPAEGSLVTEVMPVIKYRVTVCTGMVSGSGTDANVFVCLIGDQGDTGDRVLYKCINNVN KFEKGNADEFFVEAVTLKQVRRVRIGHDGKGGSSGWYLAKVIVREEGQPESEAVEFPCYRWLDKNEDDGQIVRELVPAGESPLLKNVSYHISVKTGDIPG ASSDSKVFIKLYGEKADTSKETLLVSDNDLGNYFERGRVDEFTIDTMDIGKINRILIGHDNVGLRSGWFLASVQITVPVQGRQYMFPCNRWLDKDEADGR VEVEVYPSEILPIEKLINYEVSVVTGDVRAAGTNAKVFMQIYGETGKTELIILENRSNNFERGATDIFEVEAADVGKIYKIRIGHDGKGIGDGWFLESVT LKRLATKMDETDKKKKKKKKKSEEEEEEEETKVEEVMDVYTFVAHRWLAKDEGDKELVVELVPDGESELEENTYEVHVLTGSVWGSGTDANVFLSIYGIE RGDTGERQLKRSNNLNKFEKGQVDVFTIKAIDLGELKKLRIRHDNSGSSPSWFLERVEIVDLKESTTYYFPCQRWLAVEEDDGQIVRELVPVDEAFVKKD SENDGQSLATLGLEQKAKSTTYIVKVKTGDKKNAGTDANVFITLYGSKDDTGIVSLKASKLNKNKFERGKIDEFTVESVDIGDLKKIKIGHDNAGNSNGW FLEWVEIDAPSLGQCLKFPCGRWLDKSEDDGAIERFIFPAELQTTEYIPFVPYEITVYTSDIFGAGTDADVFIVLYGSDGICTQQKSLCLNKREQRMYFE RNSVNQFIVELEDVGDIIEKIRIGHNGGGLNSGWHLDRVAIRRLLPNGKGSETITFPCERWLAKSEDDGEIIRELVPSDIFTEKLMKDGTLKQIEEEVED PLEVHTYKISVFTGDIYGAGTDANVFLNIYGDLGDTGERKLSKSETNFNKFERGQSYDGERRSLSESSVSLSSLDSSGNPKEKKSRLVKSAEEGLLIPYH ITVTTGTEYDSSTDSRVFIIIMGPQKVRTERLWLDLPEGKDEFADGSVEKFSVWGLDVGEIKKVEVGHDGATPESCWLMEELTIVVPTKGVMYNFVCKCW LARDKGDGLTSRILNILDADCVNVGIKILYEVTVVTGDIESGGTDAGIFMTVFGSNGNTEEMQLDKNGDRFERGQEDSFIMEIADIAPLRKMRIRTDAKG TRPDWFLERIVMRNLTNQEVATFTYGDWLSKVKNAKGSLVCEMPAMVNDEQMMEDTTYTIQVKTSDIGGAGTDANVSLILFGENGDSGTLALKESNKSNK FERNQMDEFNFPNMLSLGDLCKVRIWHDNKAYEIVTVTSNREDAETKENIWIILEGKLGRSKEFLMENSSKKRRFERRGSTDTFQFSSKNLGDIAAICVG HCPKDGKKSSAKADVYWHVKEIIITEMELCNKYFFRCNGKIPLRYKRRDYKVFECAKVIESFASKARSLVPVKYETIVVTGFEKGAGTDANVFITIFGLN GDSGKRALKQKFRNLFERGKTNRFYLETLDMGELKKVRIEHDNSGLAPGWLVERVEITNSATGVTTIFPCGKWLDENRGDGLTWRELFPRY* >LOXHD1_cioSav Ciona savigny approx gene model 15 PLAT domains 0 EVIVEDTKYDRTYTFPCDRWLSLYKEDCQLTRHLMAVSGRQS 1 KRTTFEITTVTGNGEKRGAGTDANVFLTMRGSKGVSPKLQLKSEYCSRRNTFERGQSDVFRVKSVNVGNLKRIRMEIDDTGFAPAWFLELQVVVVDLANPSQRIYFPCGQWLSDDEQLFRDL VGSSNPLAR 2 PKSVNAYVVHTFTGDVRGAGTDATVKITLFGDHGDSGQLVLDDSKNNFERARKDTFSLQCPHLGKIKKIRIGKGHDNGGVSPGWYLDKVVVDDTMMDCVYSFPCQRWFAK 3 DEDDGRITRELVAGVGEAGIPYQVQVFTGRFTSSCMGGRRVKSVQGKSILDGKFQRNRVDICDIESTTMLSPLDHIEIGHDNSGTGPGWYLEKVVITCPTNGCEQVFSCRKWLATDEGDGLISR QLYESKSLRK 4 QVVQSAWNCLIWTSDVRNAGTDANVFIQVYGENGKSDELPLDNETDNFETGQKDKFKINLPEIGCIYKLRVWHDDSNPFSGWHLDKIELEPIGGKKGDLYTFKCGKWLDTNEGDGEVIREIPST GPGVKR 5 PQPLVRYSVSVVTGDKRGAGTDANVSCCLYGRQGDTGNRPLTASKNNRNKFERGQLHLMLIIQTQHHRWLATDEDDGAIVRELTSEGSP 6 QLLNTSYHVAVKTGDIRGAGTDADVYIQIYGSDGDTGRLKLTRGENIRNKFERGRTDKFKLEATDIGRLERIYIGHSDSGIGAGWFLDGIEIDVPSSGMHYTFKANRWLASDEGDGKTEIELYPT QTR 7 QQDKLPYEITVATGDTLNAGTDAQVVLQLYGEDGKSEVMKLRNKTDNFERKAIDKFKIEAPDIGPITKIRIGHDGRGVGSGWFLDSVKIQRFVHKTSKRKKRRRGQNQSEVETYWFVCKRWFDKSEDDGQ 8 IIRELEYTVDVYTGDKYGAGTDSNVFLTLYGENGDSGERKLDKSVNINKFERNQVDSFKLKAADLSTLKKLKIRHDNSEIGSSWFVDRVEVKVKGETWVFPCQRWLSTKEDDGQIERTLVPV DATYQRLQQQKQGRKASIAVRDAIALET 9 KAQLTTYEVSVKTGDVWGSGTDANVYLTLFGEKDDSGINLKTSKTYRDKFERNHIDVFTVESADLGVLKKIKIGHDNAGLSPGWYLEYVEVDAASLGMKWRFPCGRWIAKDKDDGKLEKELHLAT DETVG 10 YAAKVPYEITVYTSEISGAGTDADVYVVLYGREGIATEQTSLCSSKDERKSRFNKKGIDTFVLELDDIGEVIEKLRIGHDGKGWGAGWHLDKVEVRRLLEGVKGSETSTFTCGRWLARDEDDGEIVRELVPT KIVKEQVTKSGKLKRSESKLD 11 DALKQQYKVHVFTGDVFRGGTDANVFITIFGENGDTGERKLLKSETNTDKFERGKEDIFKLDAVDLGKLFKVHIRHDNALIQPAWFLDRVEIEDTDTGSIYVFHCERWLAKNKEDGKIARSLYVV GYDGETSTRRSLATTKSGGSLGKKSILSDTIP 12 EGPAIPYTVKIETGSEEGSSCKAPAFIKIFGPSKKGTVSRRIPLEPYSGKFKRGTIETFNIEAIDVGSIKKIEIGHSSVDEEWFVESVMVNQPTLGKSYTFKCHKWFARSKDDGLVSRIFTTD DAEKLS 13 FKAKIPYEITLHTGNVDNAGTDCTITLSVYGSKGNSPDITVKKSEEDGVLLERGSIDTINVELDEIAPLKMVRVGHDGRGQRADWFCDKKTLSRELSASVGGRS 14 VLKKTNYKINVKTSNLRSAGTNANVSCIVFGTFGETGELKLKVSETHKNKFERDAMDTFTFQDKLSVGEPQKLRVWHDSKGFSSGWHLDHIEVCDESTQQKYMFPCGRWLADDEDDKQTIREL TCSNLDQ 15 AKEGGQYTATVRTSDKRDAAPNSDGWVILHGKMGKSRRMKLREGKLGRGKASTFKLDSNDLGALQKITLGLEGDNMGRWHVDDVVIENPSGGQFKFDCHAWVSNEDGTNTRGSNFDC TATKGRTISMS 16 SLLPVKYEVTVVTADEKGSGTDSNVSLTIYGTNGDSGKRPLKQRFRDLFERKQTDKFTLEILDLGDLTKVQIRLEHDDAGFNADWLCDNVTIRNPVSGQSWTFICREWIGLKKANQLFREVLPR >LOXHD1_cioInt Ciona intestinalis XM_002122635 defective assembly; frag gene model 16 PLAT domains EVIVEDTKYDRTYTFPCDRWLSLYKEDCQLTRHLMAVSGRQSKRTTFEITTVTGNF GEKRGAGTDANVFITLRGSKGVSPKLQLKSD 21 RHNTFERGNSDVFRLKSANVGNLKRVRVEIDENGFAPAWFLERVVVVDMSKPSQRIFFPCGQWLSDDEQLFRDLVGSTDPLARPKM NAYVVHVFTGDMRGAGTDAEVFITLYGGKKGQLSSGKIVLDGKFQRNRVDICDVESASMLSPLDHIEIGHDNSGSGPGWFLDKVVVTCPTNGCEQVFSCRKWFATDEGDGLTSRELYESKSLRKQVMQK SAWNCLIWTSDVRNAGTDANVFIQVYGENGKSDEIALDNETDNFETGQKDKFKINIPEVGRMYKLRVWHDDSNPFSGWHLDKIVLEPVGGKKGSSYTFNCQKWLDTNEGDGQIIREIPATGPGVKRPQPL VRYQVTVVTGDKRNAGTDANVFCCLYGRQGDTGNRPLEASKSNRNKFERGQSDEFTIEAVDLKSVNKVKIGHDDSGSGWHLKEIIVRDETNGESTVFPCDRWLATDEDDGAIVRELTSEGSPQLLNT TSYHVSVKTGDMRGAGTDADVYIQIYGSDGDGRLKLHQGENIRNKFERGRTDRFKLEATDIGRIERIYIGHNDSGVGAGWFLDGVEIDVPSSGMHYTFKSNRWLASDEGDGKTEVELYPTQTRQQDKLL PYEITVTTGDIMNGGTDAQVVVQLYGEDGKSEVIRLRNKTDNFERKAVDKFKIEAPDIGPITKIRIGHDGRGVGSGWFLDSVKIQRFVRKTSKRKKRRRESMVSNDQNEVETYWFVCRRWFDKSEDDGQIIRELVPTDESGKPIDEALQE LEYSVDVYTGDKYGAGTDSNVFITLYGENGDSGERKLDKSDNINKFERNQVDGFKLKAADLSNLKKLKIRHDNSELGSSWFVDRVEVKVKGQTWVFPCQRWLSTNEDDGQIERTLVPVDVATYQRLQQKQGSGRKASVAVRDAIALETKAQL TTYEVSVKTGDIRGSGTDANVYLVLFGENDDSGMINLKTSKTHRDKFERNHTDVFTVESADLGNLKKVKIGHDNAGLSPGWYLEYVEVDAPSLGMKWRFPSGRWIAKDKDDGKLEKELFLAMDETVGYAAK VPYEITVYTSEVSGAGTDADVYVVLYGRDGMSTEKTSLCPTKDERKSKFNKKSVDTFVLELDDVGEVIEKLRIGHDGKGWGAGWHLDKVEIRRLLEGAKGSETYSFTCGRWLARDEDDGEIVRELVPTQLVQEKVTKS QQYKVHVYTGDVFNAGTDANVYITIYGENGDTGERKLLKSETNTDKFERGKEDIFKIDAVDLGKLFKVHIRHDNALLNPSWNLNRVEVEDTDTGNRYVFHCERWLAKGKDDGKIVHVYTGDVF IPYTVKIETGSDEGSSSTASAFIKIYGPSKKGKVSRRITLDPPSGKFKKGTVESFNIDAIDVGTIKKIEIGHTGVDDEWFIESVMINQPTLGKSFTFKCHKWLARSKDDGLTTRIFTIDDAEKLTYKAK IPYEVTIHTGNVPNAGTDCLITMSVYGTKGNAPDITIKKSEEEGILLERGSVDVFNVEMEEIAPLKMIRISHDGRGQRQDWFCDKIELRNLNSGKVTVFPCDSWLSKKLGEKTLSRELSASVGGRSVLKK TTYKLNVKTSNMRSAGTNANVSCIVFGTFGDTGELRLKESETHKNKFERDSLDVFTYQDKLSIGQLQKLRVWHDSKGFSSGWHLEYIEVVDESTQQKFMFPCNRWLADDEDDKQTIRELTCSNLDQATEGHTV GQYTATITTGDKRDAASSCDGWIILHGKMGKSRRLKIRSGKMGRGKKSEVKLDAGDVGKMEAITLGLHDMGRWYVEDVTIETGNGGQYKFDCHAWVSNEDGTNTRGRTFECSATKGRTVSMSSLIP IKYEVTVVTADEKGAGTDANVKLTIYGENGDSGSRALKQRFRDLFERKQTDKFTLEILDLGELTKIRLEHDDGGFNADWLCDHVIVRNPVIGQSWTFVCREWIGTKKAGQLFREVMPRSTG* >Schistosoma mansoni (flatworm) XP_002576380 2301aa Bilatera|Platyhelminthes blast 0 MNLEDSTNDDYLKNSKSRFSFHRKEEKEHKGIICCGHNTCHLDRYNLISSVKPFTARPYTLYKVVLTTADVPGAGTSAQ 1 2 IYITLKGEWGSSTRQKLRKEKVPTRNLRFYFYPGSTNTFSVVSPDLGGLHSVFIE 0 0 HDSLRKSDSWLLESVQVFHPLTKKRYMFMCNHWFSLYKEDGLIARELFGIRSAKT 1 2 KYSIVTVTGDQEGSGTNSKVYITIYGRTGITPRIELSQENKSTGKDILCAPFGRGTSTKFIVKAPNVGAITNIRIKQDESGNEPHWFLERVVVTDMSYPQWTYYFHLSCWLSSKYGDGKSCRLVRGYREPTGTGV 1 2 ETEYKLTFYTSDQKDAGTTGEVYVKLYGEKDSSREIWVNSINQKNRRQPTSYHFTRGSTVEVYLPPCPQLGEITKLKVGHNRAGSSPSWFLNK 0 0 VIVDDLRMNRVFEFPCYAWITKPSETIIVCQKSIKKEEKEIIWQ 1 2 KIPLEIRLYTGDVPNAGTTAKVYL 12 HQKTTSENKEDVYTTPYIWLEDGNYDRNGVTLFSIDLPVPKFISPLSQLVIGHDNTGHSPSWFFDK 0 0 VQIYCPLNGVEQTFLYRKWLTSSKPGMKVEQILCEEKSLRKCTDK 1 2 KIPWEISVKTSPIANSYVTAAVSIIIFGSKDKTMKIQLDRSNLEVSSEWTKVSGNEVVEMFRPNVESKFRIYIKDIGIPCKLRIQHDNKGSNPNWHLQE 0 0 IVLTNLRTHEQYEFYCNRWLSTREDDGSILREIPAKGPGITNPLP 1 2 LYHYIIQIFTGNKPNSGTNANIFINIFGEKGDCGERWLGRSVNRNSELFQQNQ 00 MDEFVIEAVQLGSINKICIGHEERSPGYGWYLAKIVLTIKENPKYKLTFECYR 21 WFDVGEDDGQIVRELFAHSSLS 1 2 AIAYNVTVLTGTCRNAGTVANVFVHLYGLQGESKDMQLKHKETEITKFEAGKSEEFILACGKLGE 0 0 IKKIKIWHDSIAPHSGWFLDEVVVIDYFLGRRYVFHVNRWLAKNEADGLISLDLQPTSIINIER 1 2 KSSYEVIVKTGGRKYAGTDSHVYITMFGINSESKEYHLANSKTHNNKFEQNHEDLFN 0 0 LNAVGLGSLKKIRVRHTNTGIAPGWFLDYILIRESIEQKKMEYFFPCYQWLSATRLDGLVIRELSVASENIFERWKSGHDISDDSRLILE 1 2 DQSTIYHVRIHTGSQSNISSDANVQIQLYGDKDFTGNIRLYKAYYGKEDILVNKFQAGQIANFIVKAINIGNIKKIR 2 1 IEHDTAVGSARWFVERVEVEARKLGLLWKFECNRFITPQQPELILYPEPKDISFVKP 1 2 MTNYQIKTFTSDISGAETTSSVYIQLYGNDSLPSSIRRLHQNNDSEQRFQRNKIDTFYVELEELQEPFSKLRIWHNDKGSSTDWHLNKVEIRKIKTNQY 0 0 VFVTYIFPCNKWLSRNMDQAALERELIPSHMLQDQNGVLQFERQLSPKWLMNTYEVRITTGDKAYAGTDASVYITLFGENGDSGERKLTKSLTHRNKFERGQ 0 0 TDVFQLEIVDLGKINKVRIRHDNSGVNPSWYLSTIEVFNISKSQSVHGLKDQPNETHHLVKYIFNCEAWLSTEHDEKVLDRFFSAEIIKPQDLPGIVPKTSKDIVPHKLIQDLSQGKKALE 1 2 TIPYLITVITGSDRKAGTPGPVWISCVDKDKMNSEKFILCDCYNRTMLKRGTTRYFRFAGVKLNDLTEIQ 0 0 VGNDAPESPSMGWYIQSLYLSFPTIGKMYLFDCKEWLSTNRGNKKRTCNLKISNTNIVKFKP 1 2 KITYHLKITTANVKRAGTDCSINLQIFGTNGVTNCYILEKTSNRFAQGITDNISLEMEDVGKLLKMRIGHDNQ 0 0 GKNKHWNLSCVEVTVANTNQLYRFVYDDWLSLTYGKRKSLWADLPAMIGDTVQLK 1 2 ETCLDIFVKTGNMPASSTDANVYVQLFGEYGDSGEILLKQTVSNQKPFQNNS 0 0 IDHFKIPSILKLGNLARCRIWHDNKGSSPNWYCEWLEVKEVLIPGEKNLACNWKFAFNKWLSVSDDNKQLLRDAPCSEVYMNDSKGHRTIDQESIETLLTTADSMNISDLKDNPEGKL 1 2 VYEVVIETGNLKDSGTTCDAWIILEGKHGRSPKLELVNQVGNPILQINQMNTFQ 2 1 LPSFPLGDLETIRLGIQERNINKQTNPNDIQSQKWFCEKVSIKDPVSKRTYIFTIHQWLSVTPTSNLKKDVLVNYSKIIEDPYKKAFNELK 1 2 NKQTVMYKVSIYTGTKGCANTDANIFITMFSTTPGLNSGRIALKRENNNLFDRKQLDEFYVESIDL 1 2 ESIQRIIIEHDNTGVSPDWYLDKVLITNQTNNQIHLFQCYQWISKKKGDCRLWKELLVSN* 0 >LOXHD1_triAdh Trichoplax adhaerens (trichoplax) Metazoa XM_002107971 17 PLAT domains blast 0 MSASLAMQELMSDPSGHFAPKPPPGRSKINRKRPQT 1 2 APGRQITTASKAPKLWRSSTYNEPNLKRRKFIVTLNHKKDISSISRNAPYANRAYTTAMATAARTYSNALNKGQQKEIIPIYNPLCDPHLNDYYARKFGLLNSRD 0 0 ESRKNRKQ 1 2 AGVAYQFGVKTGDKKGSDTDA 2 1 VYIQVIGTKDKIPKKRLFKKQETEKTERGNLFKFDKSTVEKFAVQHRDIGDPVKLIVE 0 0 HDGNEKRHGWFLEEITLTNIQSKKSWLFPCHKWLSKYEGDRKLCYELKPLAKAGKA 1 2 VYEVSVLTGDKRGAGTDANVSVTLFGKHTSSPKIQLLKS 2 1 SKHKNPFERNNTDEFKIRTRDVGKLSKIRIEHDNAGFGPGWFLDK 0 0 VIICNLEKPNVKYYCPCNQWLAKDVGDKSISRDLTAYTDPNAAPS 1 2 AYVYIVHTFTGNKRGAGTDANVYAVIFGDSGDTGEKRLDNSKNNFEKSR 2 1 KDTFKLSCSCVGKLERLRIRHDNTGLFAGWYLDKV 1 2 VVEDPQEQQSYTFYCRRWLSKTEDDGEICRDLIVSASGDDGDDSVAPK 1 2 GYPYHIHVTTSDVKNAGTDAEVYVVMHGEGKKSKELNSGKLVLANSEKKKNTFERAMTDIFHMECAEMLSPLTKLTVGHDNKGLAAGWHLDR 0 0 IVIDCPTTGIEQTFLCQQWLDRKAGDGLTERELVEAFDMRKTRRP 1 2 KQLWFAWIWTSDIRGAGTDANVSMQIYGDKGKSQEIKLGNNTDNFEQATLDKFK 0 0 LEIDQVGVPYKLRIGHDNSNAFPGWHLDKVKLENMNDKEQYLFNCNRWLSRSEEDNEIIRELPASGPNCPNYP 1 2 IVIYEVSVHTGNKMGGGTDANVFIKIYGELGDSGYRPLKSSKSHNNKFERNQVDVFHIEAVTLKALKKIKIGHDGNNP 1 2 GAGWFLDKVVIKELNGEASNEFPCNR 2 1 WLSKSEDDGQIVRELFLKSDTPLLK 1 2 TTSYHISVKTGDVRNAGTDANVFIQIFGAKDDTGRVRLKQSLNTSNKFERNRIDKFIIEAAQIGK 0 0 IEKIIIGHDGKGLGSGWFLDYIELDVPSVGRLYRFSCHQWFDSTEGDRKVERELYPSECIKSAA 1 2 KIPYQISVHTGDIRHAGTDSNVFAVIYGENGKTEELKLRNKSDNFERGQVDVFK 0 0 VECEDVGKLRKLRIGHDSAGMGSAWFLDK 0 0 VYVRRLPPKSGKKSKETDEREEETAKKDADEPEKLDENNYLFVANRWLSKEEGDRQTVIEISPVGVDGALA 1 2 EMTYTIRVITGNKFGCGTNANVFINMYGEEGDSGERQLKKSETHTDKFERNQ 0 0 EDVFKISCLSLGELKKIKIRHDNSGFRPAWFLDKVIIEVGESKYQFMCDRWLAKDEDDGQISRELLPQSDEQSRAEAIGASKDLQKK 1 2 VASTTYNVSVTTGDIKGAGTDANVHIVLYGEKDDTGLIHLKNSTTHSNKFERNQEDRFVVEAIDIGELKKIK 2 1 IGHDNKGGMAGWFLNKVEIDIPSLGRRLLFPCGRWIDKGKDDGALERELYPLNEAEETYRP 1 2 HIPYEVTVYTTDKRGASTGANVYVVIYGEENQTEQASLEPDKKRRKQYFKNGAIDKFVLE 0 0 LDDVGEEITKLRIGHDGKGWGAGWHLDKVEICRLLDGGKASKKFTFQCNRWLASDEDDGAIVRELVPSEIVEKSSKDGGQVKTKVTKPTDGLK 1 2 VKPYTIHVFTGDVDGAGTNANVFLTIFGESGDSGERKLAKSDTHYDKFERNQ 0 0 EDIFHIEAADLGRLFKVKIRHDNTGSLFSPAWFLNRIEIVDDENEETTAFPCERWLAKKKDDGKIDRTLFVKGWEGDTSSVATTRSK 1 2 ASFRSNSTDLGEMVSASGRRASSRKGPVMAPIDEK 1 2 CVPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVS 1 2 HNSYMPDSGIYLKEIEVDVPTIGNKYIFPCNRWLAKDKDDSKTSRIFTAANAQVTSYTKPYELTIHTGDVQNA 1 2 GTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIP 0 0 LELDDVGTIKKIRIGHDGKGSRTDWYLEK 0 0 ASIQRMDTLDMYMFRANQWFSKKIDDKKLVREIPAETSKEGATTIK 1 2 KINYVLSTHTSDKRGSGTDANVFVIIFGENGDSGEIALKKSETNWNKFEKGQTDVFLINDRLSLGRLQKLRIWHDNA 1 2 GFGASWHLASVDIVDESTGVKYTFPCDKWLSKSNGDKLILRELPCAETAGNTAASKATKQESKSGKA 1 2 EYEIAFTTGTEKRAGTNQDVAIVLKGKSEKSREFLIENNEDKKYFSKGKTNKFTYTCKPLGDITKAIVSHRESAIGEEPDSKNSSWYLKVVTVVHKASGTT 2 1 YKFPCNKWIDLDDDEENNSSVTLKCKSADVAASSKAKPV 1 2 ELKPVKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* 0 >LOXHD1_triAdh Trichoplax adhaerens (trichoplax) Metazoa XM_002107971 17 PLAT domains blast MSASLAMQELMSDPSGHFAPKPPPGRSKINRKRPQTAPGRQITTASKAPKLWRSSTYNEPNLKRRKFIVTLNHKKDISSISRNAPYANRAYTTAMATAARTYSNALNKGQQKEIIPIYNPLCDPHLNDYYARKFGLLNSRDESRKNRKQAG 01 VAYQFGVKTGDKKGSDTDAVYIQVIGTKDKIPKKRLFKKQETEKTERGNLFKFDKSTVEKFAVQHRDIGDPVKLIVEHDGNEKRHGWFLEEITLTNIQSKKSWLFPCHKWLSKYEGDRKLCYE LKPLAKAGK 02 AVYEVSVLTGDKRGAGTDANVSVTLFGKHTSSPKIQLLKSSKHKNPFERNNTDEFKIRTRDVGKLSKIRIEHDNAGFGPGWFLDKVIICNLEKPNVKYYCPCNQWLAKDVGDKSISRD LTAYTDPNAAPSA 03 YVYIVHTFTGNKRGAGTDANVYAVIFGDSGDTGEKRLDNSKNNFEKSRKDTFKLSCSCVGKLERLRIRHDNTGLFAGWYLDKVVVEDPQEQQSYTFYCRRWLSKTEDDGEICRDLI VSASGDDGDDSVAPKG 04 YPYHIHVTTSDVKNAGTDAEVYVVMHGEGKKSKELNSGKLVLANSEKKKNTFERAMTDIFHMECAEMLSPLTKLTVGHDNKGLAAGWHLDRaIVIDCPTTGIEQTFLCQQWLDRKAGDGLTERELVEA FDMRKTRRPK 05 QLWFAWIWTSDIRGAGTDANVSMQIYGDKGKSQEIKLGNNTDNFEQATLDKFKLEIDQVGVPYKLRIGHDNSNAFPGWHLDKVKLENMNDKEQYLFNCNRWLSRSEEDNEIIRELPAS GPNCPNYPI 06 VIYEVSVHTGNKMGGGTDANVFIKIYGELGDSGYRPLKSSKSHNNKFERNQVDVFHIEAVTLKALKKIKIGHDGNNPGAGWFLDKVVIKELNGEASNEFPCNRWLSKSEDDGQIVRELFLK SDTPLLKT 07 TSYHISVKTGDVRNAGTDANVFIQIFGAKDDTGRVRLKQSLNTSNKFERNRIDKFIIEAAQIGKIEKIIIGHDGKGLGSGWFLDYIELDVPSVGRLYRFSCHQWFDSTEGDRKVERELYPS ECIKSAAK 08 IPYQISVHTGDIRHAGTDSNVFAVIYGENGKTEELKLRNKSDNFERGQVDVFKVECEDVGKLRKLRIGHDSAGMGSAWFLDKVYVRRLPPKSGKKSKETDEREEETAKKDADEPEKLDENNYLFVANRWLSKEEGDRQTVIEIS PVGVDGALAE 09 MTYTIRVITGNKFGCGTNANVFINMYGEEGDSGERQLKKSETHTDKFERNQEDVFKISCLSLGELKKIKIRHDNSGFRPAWFLDKVIIEVGESKYQFMCDRWLAKDEDDGQISRELLPQ SDEQSRAEAIGASKDLQKKVAS 10 TTYNVSVTTGDIKGAGTDANVHIVLYGEKDDTGLIHLKNSTTHSNKFERNQEDRFVVEAIDIGELKKIKIGHDNKGGMAGWFLNKVEIDIPSLGRRLLFPCGRWIDKGKDDGALER ELYPLNEAEETYRPH 11 IPYEVTVYTTDKRGASTGANVYVVIYGEENQTEQASLEPDKKRRKQYFKNGAIDKFVLELDDVGEEITKLRIGHDGKGWGAGWHLDKVEICRLLDGGKASKKFTFQCNRWLASDEDDGAIVRELVPS EIVEKSSKDGGQVKTKVTKPTDGLKV 12 KPYTIHVFTGDVDGAGTNANVFLTIFGESGDSGERKLAKSDTHYDKFERNQEDIFHIEAADLGRLFKVKIRHDNTGSLFSPAWFLNRIEIVDDENEETTAFPCERWLAKKKDDGKIDRTLFVK GWEGDTSSVATTRSKASFRSNSTDLGEMVSASGRRASSRKGPVMAPIDEKC 13 VPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVSHNSYMPDSGIYLKEIEVDVPTIGNKYIFPCNRWLAKDKDDSKTSRIFTAA 14 NAQVTSYTAGTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIPLELDDVGTIKKIRIGHDGKGSRTDWYLEKASIQRMDTLDMYMFRANQWFSKKIDDKKLVREI PAETSKEGATTIKK 15 INYVLSTHTSDKRGSGTDANVFVIIFGENGDSGEIALKKSETNWNKFEKGQTDVFLINDRLSLGRLQKLRIWHDNAGFGASWHLASVDIVDESTGVKYTFPCDKWLSKSNGDKLILREL PCAETAGNTAASKATKQESKSGK 16 AEYEIAFTTGTEKRAGTNQDVAIVLKGKSEKSREFLIENNEDKKYFSKGKTNKFTYTCKPLGDITKAIVSHRESAIGEEPDSKNSSWYLKVVTVVHKASGTTYKFPCNKWIDLDDDEENNSSVTLKC KSADVAASSKAKPVELKP 17 VKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* >Monosiga brevicollis (monosiga) Eukaryota|Choanoflagellates XM_001742822 17 PLAT domains blast exons colored; PLAT domains numbered MADLPVPTTALQRLQLERSRYSFAAADPPPSFVQRYTSADVLSHRLYAQGLASDVGPGSHSSLGMSRRVELPPYNPLNDPALANYFARKFEWNSTRSVGAGSGSRRSASASLTRTRQPVRGASAHGHTKSRTKSSHRK 01 GHASLEIFTSKRSNPVSKSEKYITVVGTKGRSDAIQLAAPGVHFRAGNKDVFQVNLTGIGKPTKVILENTGTKRTDGWCVSKVVLVKKTDKGTRRYRFSGPVWLSKHHDEMKLKR VLHIDDEDIGSG 02 NGYRIDCYTGDVANAGTDAVATIQLFGSKGQSPMVELRRSDGQAFQRARVATFTLDDLANLGKLKKLVISHNGHGMASGWFLDKIIVTSLSSNKATVFPCDAWLDRKNGRSKELVAR TAAAGAEGM 03 TTFTIRVMTGDRRGAGTDANVQCTLFGEDGESGPHTLNTSRNDFRRGHTDVFAVSSRKIGTLKRLRIWHDNGGAGPAWFLDAVEVVDEASGQTYRFECNRWLAKDEDDGQISRELTCN GDSSWGL 04 KSYKLTIFTGDKRNAGTSANVFCKLVGERGASDNVILENSSKNFQRDRTDIFTVEASDLGSLRHIVLGHDNHGMGAGWYVERFSLEVPSEGKLYNVDVKQWFATDMSDGAIERTFRLD NAETLDIAQR 05 LDWKCTIYTSDVANAGTDANVFMQVYGKKGKTDVVPLKNKSDTFERGQTDELRVQLINVGSLRKLRVWHDNKGMASGWHLDRIVLSRDGEEYIFPCAEWLAVSEGDKEIVRELPAT GPNVKKPLQL 06 VEYTVRVATGHARFAGTNADVFVMLTGELGDSGKRALLRSQTNRNKFERGKEDVFTVAAVDLGKLTSVTVGHNNAGTSAGWFLDKIVVLDPRRGEEEEFPCHRWLAVDADDGQIERELVPK MAEHQAAAT 07 TTYIVKIKTGDVRHAGTDANVFVQLFGKTGESTQLKLRNSETYSDAFERNKMDIFKFELLDLGDLSRILVGHDNKGMGAAWFLDYVEVEVPSIRTRWKFPCSRWFSKSQDDGLTER EIYAEKEAGEPMEEDVS 08 APYLFRFYTSDVAFAGTDANVSVVLYGDEGKTEELVVNNQSDNFERGKADDFKLACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQARIYTFEHNDWLRKGTKAKPFMVEL PLRRIETVDDNGREVVEELALDANK 09 RTYRVKVHTGDQKGAGTDANVYVNLHGSLGDSGDRHLKNSLTHTNKFQRKTVDEFDIDAVTLGDINKVKVWHDNAGLGAAWYLEKIEVVDTADDKTYIFPCAQWFAKSMGDGQIAREL GVLEEQKPADFQTKNVG 10 FKYRISVHTSDVKHAGTDANVDIVLYGEKGDTGKIRLAKSETHRDMWERGNCDVFTVSAIELGDLKRVDIMHDGKGVGSGWHLNKVVVDAPQAGKTWTFMCDAWLDKATDDGTMAK TLYASADAIEEYSAH 11 VPYEIIIKTSDVRNAGTDANVFIDLYGRDQEERDLTAHHEFKDAVKAHFERNLEDRFNVELPDVGSIYKIRLGHDGKGMSSSWHVASVVVINQRTHERFEFPCDAWLSKDKDDKKLVREFAVG EVKALEGDKVVRRESILSLQE 12 AIYKIHVFTGDIKHAGTDANIYVQIFGDTGDSGEIKLEKSETYRDKFERGHEDIFTHRCLDLGPLRKIKVRSDGKGLMGGDWYLDRVEVHQENDLSEPPVRFVCQDWFKRGKQEGDTLEREITAQ VDAAVAMKEATALEDEAARLQRKATTLSRKSTKGSPLRKGTGTTETNAELEEAQRQANIARRKADEAKERAGLAGADDTQ 13 LEYKVTVYTGTDTQAGTTANVWLQLFGEKDAPLTPAPPSPSKLSRSGSLFGRRRRASSDAQSVSSSLSAGASGPRETSTGRLQLNNAPADLQSGAKTTFTVTGLDVGELVGLEIGHDDERDKWYLEQVVVEVPKSATHAARRYEFKAGVWLAARADGSTSGSAK GKSKASVRLQPSEIHAGSERI 14 LEYTLKVYTASADGAGCTAVPQVQLFGDKHTTEALPLRAGGDILPASVVETQHRVPDLGALLKVRLMVPRGSSWTVEKVEFGRAGQTPITFVGSDGSAVTLGAERLS FDFLPAAAPASSGGKGKRRGSTTSSVAVAQQ 15 TSYRVYVTTADERGTGTDANVSIILYGAMGDSGEHSLTKSETFDDPFERGNTDVFTLEVPDLGELQRARIWHDGKGMFSSWKLDKIVVVVEATQSRYELPCGQWLSKNKGDKQLTRDLAVASKR VNALT 16 GTYKLEVATDSRAGGGCKGPVRIMLLDAEKNQLPLTLEPPGGEFAPGSVEHLVFDNVMLLGPLTELRIRRKPSGASSRRGAEDDDEDDNEASGASSQSGSAVSPWHLEHIIVKHLQSGQSFVFKGPSKGLSRSRAKLSVHTEA TTEEAVAQATKASV 17 AQYEVAVTTGTERGAGTDSNVFVTLFGKNGDSGERALAKSKTFRNMFESGNTDVFDVECQDLGELTKIEVKSDLKGFGAAWQLDKIKVTRTGSQNSWQFKCDQWFDKKQGAEHTFSVA S*
>LOXHD1_homSap_full dna: each line is an exon ATGATGCCCCAGAAGAAAAGGCGGAGGAAGAAGGACATCGACTTCCTGGCCCTGTACGAAGCGGAGCTGCTGAACTACGCCTCGGAGGACGACGAGGGGGAGCTGGAACACGAGTACTACAAGGCCAGAG TGTATGAAGTGGTCACAGCCACGGGGGATGTTCGCGGTGCAGGGACGGATGCCAATGTCTTCATCACGCTTTTTGGAGAGAATGGGCTCTCTCCCAAGCTCCAGCTCACCAGCAA GAGCAAGTCTGCCTTTGAGAAGGGCAACGTCGATGTGTTCCGGGTGAGAACCAACAATGTGGGCCTCATCTATAAAGTCAG GATTGAGCATGACAACACGGGCTTGAATGCCAGCTGGTACCTGGACCATGTGATTGTGACCGACATGAAGAGGCCTCATCTCCGTTACTACTTCAACTGCAACAACTGGCTGAGCAAGGTGGAAGGTGACCGCCAGTGGTGCCGTGACCTGCTGGCCAGCTTCAACCCCATGGACATGCCCAGAG GTAATAAGTATGAAGTCAAGGTATACACTGGTGATGTAATTGGTGCAGGGACAGATGCTGATGTCTTCATCAATATTTTTGGAGAGTATGGAGACACAG GGGAGCGTAGGCTAGAAAATGAAAAGGACAACTTTGAAAAGGGAGCTGAAGACAGGTTCATCCTGGATGCCCCGGATTTGGGGCAGCTGATGAAGATCAATGTTGGCCACAACAATAAGGGGGGCTCTGCAGGTTGGTTCCTGTCCCAG ATAGTCATTGAAGATATTGGGAACAAAAGAAAATATGACTTCCCCCTTAACCGCTGGCTGGCCTTGGACGAAGACGATGGCAAAATCCAAAGGGATATCTTAGTGGGCGGAGCTGAGACCACAG CTATTACGTATATTGTCACCGTCTTCACTGGGGATGTCCGGGGGGCTGGTACCAAATCCAAAATCTACTTGGTCATGTATGGGGCCAGAGGGAATAAGAACAGTGGGAAAATCTTCCTGGAGGGCGGCGTGTTTGACCGAGGCCGCACGGACATCTTCCACATCGAGCTGGCTGTCCTCCTTAGCCCCCTGAGTCGGGTCTCCGTCGGGCATGGCAATGTGGGTGTCAACAGAGGCTGGTTCTGTGAGAAG GTGGTGATTCTGTGCCCCTTCACTGGTATCCAGCAGACCTTCCCTTGTAGCAACTGGCTGGATGAGAAGAAAGCGGATGGGTTGATTGAGAGGCAGCTCTATGAGATGGTGTCTCTCAGGAAGAAGCGGCTGAAAA AATTCCCTTGGTCCCTGTGGGTCTGGACAACCGACCTAAAGAAAGCTGGTACCAACTCTCCCATCTTCATCCAGATTTATGGGCAGAAGGGGCGGACAGATGAGATTCTCCTGAATCCCAACAACAAGTGGTTCAAACCCGGCATAATCGAGAAGTTTAGG ATTGAGCTCCCGGATCTTGGCAGGTTTTATAAGATTCGAGTATGGCATGATAAAAGGAGTTCTGGTTCTGGATGGCATTTAGAAAGG ATGACCCTGATGAACACTCTGAACAAAGACAAGTACAACTTCAATTGCAACCGCTGGCTGGATGCCAATGAGGATGACAATGAGATAGTGAGGGAAATGACTGCAGAAGGCCCAACAGTGCGCAGGATCATGGGCA TGGCCCGGTACCATGTGACTGTGTGCACAGGTGAACTTGAAGGTGCTGGGACCGATGCCAACGTCTATCTCTGCCTTTTTGGTGATGTGGGGGACACGGGGGAACGGCTGCTCTACAACTGCAGGAATAACACAGACCTGTTTGAAAAGGGCAAT GCTGACGAGTTCACTATCGAGTCTGTCACCATGCGGAATGTGAGGCGGGTGAGGATCAGACACGATGGCAAAGGCTCCGGCAGCGGCTGGTACCTGGACAGAGTGCTGGTGAGAGAGGAGGGGCAGCCTGAGAGCGACAACGTGGAGTTCCCATGTCTCAG GTGGTTGGACAAGGATAAGGATGATGGGCAGCTGGTCCGAGAGTTGCTACCCAGTGACAGCAGCGCGACACTGAAGA ACTTTCGCTATCACATCAGCTTGAAGACTGGGGATGTCTCTGGGGCCAGCACGGATTCTAGAGTCTACATCAAGCTCTATGGGGATAAATCTGACACCATCAAGCAAGTTCTTCTTGTCTCTGACAACAACCTCAAAGACTACTTTGAACGTGGCCGGGTGGATGAGTTCACCCTCGAGACCCTGAACATTGGAAAT ATCAACCGGCTGGTGATTGGGCATGACAGCACTGGCATGCATGCCAGCTGGTTCCTGGGCAGCGTTCAGATCCGTGTGCCCCGTCAAGGCAAGCAGTACACCTTTCCCGCCAACCGCTGGCTGGACAAGAACCAGGCTGACGGGCGCCTGGAGGTGGAGCTGTATCCCAGCGAGGTGGTGGAGATCCAGAAAT TGGTCCACTATGAGGTTGAGATTTGGACAGGAGATGTGGGTGGCGCAGGCACCAGTGCCCGAGTCTACATGCAGATCTATGGAGAGAAAGGCAAGACAGAAGTGCTCTTCCTCTCCAGCCGCTCAAAAGTTTTTGAACGGGCGTCCAAGGACACATTCCAG CTTGAGGCGGCCGACGTGGGCGAGGTCTATAAGCTCCGGCTCGGGCACACGGGCGAGGGCTTTGGGCCCAGCTGGTTCGTGGACACCGTGTGGCTGCGGCACCTGGTGGTGCGGGAGGTGGACCTCACGCCGGAGGAGGAGGCCCGGAAGAAGAAGGAGAAGGACAAGCTGCGGCAGCTGCTCAAGAAGGAGCGGCTGAAGGCCAAGCTGCAGAGGAAGAAGAAGAAGAGGAAGGGCAGCGACGAAGAGGACGAGGGGGAGGAAGAGGAGTCGTCCTCATCAGAGGAGTCCTCGTCAGAGGAGGAGGAGATGGAAGAAGAGGAGGAAGAGGAGGAGTTTGGGCCGGGGATGCAGGAGGTGATTGAGCAGCACAAGTTCGAAGCCCACCGCTGGCTGGCCCGGGGCAAGGAGGACAACGAACTTGTCGTGGAGTTGGTGCCAGCTGGCAAGCCGGGTCCTGAGC GAAACACCTATGAGGTTCAGGTGGTCACGGGGAATGTGCCCAAGGCCGGCACTGATGCTAACGTCTACCTAACCATCTACGGCGAGGAGTATGGAGACACGGGCGAACGACCCCTGAAGAAGTCAGACAAGTCCAACAAATTTGAGCAGGGGCAG ACAGACACCTTCACCATCTATGCCATTGACCTGGGGGCCCTGACCAAGATTCGGATTCGCCACGACAACACAGGCAACAGAGCAGGCTGGTTCCTGGACAGAATAGACATTACTGACATGAACAACGAGATCAC GTACTACTTTCCATGCCAACGTTGGCTGGCAGTGGAGGAAGATGATGGCCAGCTGTCCAGGGAGCTGTTGCCAGTGGATGAGTCCTATGTGCTGCCACAGAGCGAGGAGGGTAGGGGAGGCGGTGACAACAACCCCCTCGACAACCTGGCCCTGGAGCAGAAAG ATAAATCTACCACATTCTCAGTGACCATAAAGACTGGGGTTAAGAAGAATGCGGGCACAGATGCTAATGTCTTCATCACACTCTTTGGCACACAGGATGACACTG GAATGACCCTCCTGAAGTCCTCCAAGACAAACAGCGATAAGTTTGAGAGGGACAGCATTGAAATCTTCACGGTGGAGACGCTGGATCTGGGAGACCTGTGGAAAGTCCGGCTTGGCCATGACAACACAG GCAAGGCCCCAGGCTGGTTTGTAGACTGGGTAGAGGTGGATGCCCCATCTCTTGGGAAGTGCATGACGTTTCCCTGTGGCCGCTGGCTGGCCAAAAACGAAGACGACGGGTCCATCATCAGAGACCTCTTCCATGCAGAGCTTCAGACGAGGCTGTACACACCAT TTGTTCCTTACGAGATCACCCTCTACACCAGTGATGTCTTTGCTGCTGGGACAGATGCCAACATCTTCATCATCATCTATGGCTGCGATGCCGTGTGCACCCAGCAGAAGTATCTGTGTACCAACAAGAGGGAACAGAAGCAGTTCTTTGAGAGGAAGTCTGCCTCCCGCTTCATCGTAGAG TTAGAAGATGTGGGAGAAATCATTGAAAAAATTCGGATTGGCCATAATAACACGGGCATGAATCCTGGGTGGCACTGCTCTCACGTGGACATCCGCAGGCTCCTCCCGGATAAAGAC GGTGCAGAGACCTTGACTTTCCCATGCGATCGGTGGCTTGCCACCTCTGAGGATGACAAAAAGACCATTCGAGAACTGGTTCCATATGACATCTTCACTGAGAAATACATGAAAGATGGGTCCTTACGGCAAGTCTACAAGGAAGTAGAAGAGCCTCTGGACa TTGTGCTGTACTCGGTGCAGATCTTCACAGGGAACATTCCTGGGGCAGGGACGGATGCCAAGGTGTACATCACCATCTATGGAGACCTCGGGGACACTGGGGAGCGATACCTTGGCAAGTCAGAGAACCGGACCAACAAGTTCGAGAGAGGAACG GCTGACACCTTCATCATCGAGGCCGCTGACCTAGGCGTCATCTACAAGATCAAGCTCCGCCATGACAACTCCAAGTGGTGCGCAGACTGGTACGTGGAGAAGGTGGAGATCTGGAATGACACCAACGAGGACGAGTTCCTGTTCCTATGCGGGCGCTGGCTCTCCCTGAAGAAGGAGGATGGGCGACTCGAGAGGCTCTTTTACGAGAAG GAGTACACTGGGGACCGCAGCAGCAACTGCAGCAGCCCTGCTGACTTCTGGGAGATCGCCCTGAGCTCCAAGATGGCCGATGTCGACATCAGCACAGTGACCGGGCCCATGGCTGACTACGTTCAAGAGGGCCCAA TTATTCCCTACTATGTGTCAGTCACCACTGGGAAGCACAAGGACGCGGCCACTGACAGCCGAGCCTTCATCTTTCTCATCGGGGAGGATGATGAACGTAGTAAGCGCATCTGGTTGGACTACCCCCGAGGGAAGAGGGGCTTCAGCCGTGGCTCTGTGGAGGAGTTCTACGTCGCAGGCTTGGATGTGGGCATCATCAAGAAAATAGAG CTGGGCCATGACGGGGCCTCCCCTGAGAGCTGCTGGCTGGTGGAAGAGTTGTGTTTGGCAGTGCCCACCCAGGGCACCAAGTACATGTTGAACTGTAACTGCTGGCTGGCCAAGGACAGAGGCGACGGCATCACCTCCCGTGTCTTCGACCTCTTGGATGCCATGGTGGTGAACATTGGGGTGAAG GTTCTCTATGAAATGACGGTGTGGACAGGGGATGTGGTTGGCGGGGGCACTGACTCCAACATCTTCATGACCCTCTACGGCATCAACGGGAGCACAGAGGAGATGCAGCTGGACAAAAAGAAAGCCAG GTTTGAGCGGGAGCAGAACGACACCTTCATCATGGAGATCCTAGACATTGCTCCATTCACCAAGATGCGGATCCGGATTGATGGCCTGGGCAGTCGGCCGGAGTGGTTCCTGGAGAGG ATCCTACTGAAGAACATGAACACTGGAGACCTGACCATGTTCTACTATGGAGACTGGCTGTCCCAGCGGAAGGGCAAGAAGACCCTGGTGTGTGAAATGTGTGCCGTTATCGATGAGGAAGAAATGATGGAGTGGACCTCCTACACCGTCGCAGTTAAGACCAGCGACATCCTGG GAGCAGGCACTGATGCCAACGTGTTCATCATCATCTTCGGGGAGAACGGGGATAGTGGGACACTGGCCCTGAAGCAGTCGGCAAACTGGAACAAGTTTGAGCGGAACAACACGGACACATTCAACTTCCCTGACATGCTGAGCTTGGGCCACCTCTGCAAGCTGAGGGTCTGGCACGACAACAAAG GGATATTTCCTGGCTGGCATCTGAGCTATGTCGATGTGAAGGACAACTCCCGCGACGAGACCTTCCACTTCCAGTGTGACTGCTGGCTCTCCAAGAGTGAGGGTGACGGGCAGACGGTCCGCGACTTTGCCTGTGCCAACAACAAGATCTGTGATGAGCTGGAAGAGACCA CCTACGAGATCGTCATAGAAACGGGCAACGGAGGCGAAACCAGGGAGAACGTCTGGCTCATCCTGGAGGGCAGGAAGAACCGATCCAAAGAGTTTCTCATGGAAAATTCTTCTAGGCAGCGGGCCTTTAGGAA GGGGACCACAGACACGTTTGAGTTTGACAGCATCTACTTGGGGGACATTGCCTCCCTCTGTGTGGGCCACCTTGCCAGGGAAGACCGGTTTATCCCCAAGAGAGAACTTGCCTGGCATGTCAAGACCATCACCATCACCGAGATGGAGTACGGCAATGT GTACTTCTTTAACTGTGACTGCCTCATCCCCCTCAAGAGGAAGAGGAAGTACTTCAAGGTATTCGAGGTTACCAAGACGACAGAGAGCTTTGCCAGCAAGGTCCAGAGCCTGGTGCCCGTCAAGTACGAAGTCATCGTGACAACAGGCTATGAGCCAGGGGCAGGCACTGATGCCAACGTCTTCGTGACCATCTTTGGGGCCAACGGAGACACAGGCAAGCGGGAGCTGAAGCAGAAAATGCGCAACCTCTTCGAGCGGGGCAGCACAGACCGCTTCTTCCTGGAGACGCTGGAGCTGGGTGAGCTGCGCAAGGTGCGCCTGGAGCACGACAGCAGTGGCTACTGCTCAGGCTGGCTGGTGGAGAAGGTGGAGGTCACCAACACCAGCACCGGCGTGGCCACCATCTTCAACTGTGGCAGGTGGCTGGACAAGAAGCGGGGGGATGGACTCACCTGGAGAGACCTCTTCCCTTCTGTC
Reference PLAT domain sets
>LOXHD1_homSap_PLAT:01 RVYEVVTATGDVRGAGTDANVFITLFGENGLSPKLQLTSKSKSAFEKGNVDVFRVRTNNVGLIYKVRIEHDNTGLNASWYLDHVIVTDMKRPHLRYYFNCNNWLSKVEGDRQWCRD >LOXHD1_homSap_PLAT:02 NKYEVKVYTGDVIGAGTDADVFINIFGEYGDTGERRLENEKDNFEKGAEDRFILDAPDLGQLMKINVGHNNKGGSAGWFLSQIVIEDIGNKRKYDFPLNRWLALDEDDGKIQRDILVGG >LOXHD1_homSap_PLAT:03 ITYIVTVFTGDVRGAGTKSKIYLVMYGARGNKNSGKIFLEGGVFDRGRTDIFHIELAVLLSPLSRVSVGHGNVGVNRGWFCEKVVILCPFTGIQQTFPCSNWLDEKKADGLIER >LOXHD1_homSap_PLAT:04 FPWSLWVWTTDLKKAGTNSPIFIQIYGQKGRTDEILLNPNNKWFKPGIIEKFRIELPDLGRFYKIRVWHDKRSSGSGWHLERMTLMNTLNKDKYNFNCNRWLDANEDDNEIVREM >LOXHD1_homSap_PLAT:05 ARYHVTVCTGELEGAGTDANVYLCLFGDVGDTGERLLYNCRNNTDLFEKGNADEFTIESVTMRNVRRVRIRHDGKGSGSGWYLDRVLVREEGQPESDNVEFPCLRWLDKDKDDGQLVRELLPS >LOXHD1_homSap_PLAT:06 FRYHISLKTGDVSGASTDSRVYIKLYGDKSDTIKQVLLVSDNNLKDYFERGRVDEFTLETLNIGNINRLVIGHDSTGMHASWFLGSVQIRVPRQGKQYTFPANRWLDKNQADGRLEV >LOXHD1_homSap_PLAT:07 VHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQLEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLR >LOXHD1_homSap_PLAT:08 NTYEVQVVTGNVPKAGTDANVYLTIYGEEYGDTGERPLKKSDKSNKFEQGQTDTFTIYAIDLGALTKIRIRHDNTGNRAGWFLDRIDITDMNNEITYYFPCQRWLAVEEDDGQLSRELLPV >LOXHD1_homSap_PLAT:09 TTFSVTIKTGVKKNAGTDANVFITLFGTQDDTGMTLLKSSKTNSDKFERDSIEIFTVETLDLGDLWKVRLGHDNTGKAPGWFVDWVEVDAPSLGKCMTFPCGRWLAKNEDDGSIIRDLFHA >LOXHD1_homSap_PLAT:10 VPYEITLYTSDVFAAGTDANIFIIIYGCDAVCTQQKYLCTNKREQKQFFERKSASRFIVELEDVGEIIEKIRIGHNNTGMNPGWHCSHVDIRRLLPDKDGAETLTFPCDRWLATSEDDKKTIRELVP >LOXHD1_homSap_PLAT:11 VLYSVQIFTGNIPGAGTDAKVYITIYGDLGDTGERYLGKSENRTNKFERGTADTFIIEAADLGVIYKIKLRHDNSKWCADWYVEKVEIWNDTNEDEFLFLCGRWLSLKKEDGRLERLFYEK >LOXHD1_homSap_PLAT:12 IPYYVSVTTGKHKDAATDSRAFIFLIGEDDERSKRIWLDYPRGKRGFSRGSVEEFYVAGLDVGIIKKIELGHDGASPESCWLVEELCLAVPTQGTKYMLNCNCWLAKDRGDGITSRVFD >LOXHD1_homSap_PLAT:13 VLYEMTVWTGDVVGGGTDSNIFMTLYGINGSTEEMQLDKKKARFEREQNDTFIMEILDIAPFTKMRIRIDGLGSRPEWFLERILLKNMNTGDLTMFYYGDWLSQRKGKKTLVC >LOXHD1_homSap_PLAT:14 TSYTVAVKTSDILGAGTDANVFIIIFGENGDSGTLALKQSANWNKFERNNTDTFNFPDMLSLGHLCKLRVWHDNKGIFPGWHLSYVDVKDNSRDETFHFQCDCWLSKSEGDGQTVRDFAC >LOXHD1_homSap_PLAT:15 TTYEIVIETGNGGETRENVWLILEGRKNRSKEFLMENSSRQRAFRKGTTDTFEFDSIYLGDIASLCVGHLAREDRFIPKRELAWHVKTITITEMEYGNVYFFNCDCLIPLKRKRKYFKVF >LOXHD1_homSap_PLAT:16 VKYEVIVTTGYEPGAGTDANVFVTIFGANGDTGKRELKQKMRNLFERGSTDRFFLETLELGELRKVRLEHDSSGYCSGWLVEKVEVTNTSTGVATIFNCGRWLDKKRGDGLTWRDLFPS >PKD1L1_homSap_PLAT chr7: 47780815 polycystin-1L1 GPS-TM-LH2-TM pdb:3CWZ QLYAVVIDTGFRAPARLTSKVYIVLCGDNGLSETKELSCPEKPLFERNSRHTFILSAPAQLGLLRKIRLWHDSRGPSPGWFISHVMVKELHTGQGWFFPAQCWLSAGRHDGRVERELTC >PKD1_homSap_PLAT chr16: 2078712 polycystin 1 GPS-TM-LH2-TM pdb:3CWZ FKYEILVKTGWGRGSGTTAHVGIMLYGVDSRSGHRHLDGDRAFHRNSLDIFRIATPHSLGSVWKIRVWHDNKGLSPAWFLQHVIVRDLQTARSAFFLVNDWLSVETEANGGLVEK >PKD1L2_homSap_PLAT chr16: 79691985 polycystin 1-like 2 signal...GPS-TM-LH2-TM pdb:3CWZ YHYLVTVYTGHRRGAATSSKVTVTLYGLDGEREPHHLADPDTPVFERGAVDAFLLSTLFPLGELRSLRLWHDNSGDRPSWYVSRVLVYDLVMDRKWYFLCNSWLSINVGDCVLDKVFPVA >PKD1L3_homSap chr16:70,556,160 GPS-TM-LH2-TM polycystic kidney disease 1-like 3 FHYLIQVYTGYRRSAATTAKVVITLYGSEGRSEPHHLCDPQKTVFERGGLDVFLLTTWTSLGNLHSLRLWHDNSGVSPSWYVSQVIVCDMAVKRKWHFLCNCWLAVDLGDCELDRVFIPV >PKDREJ_homSap_PLAT chr22: 45030224 receptor for egg jelly-like signal...GPS-TM-LH2-TM pdb:3CWZ CYLVTIFTGSRWGSGTRANVFVQLRGTVSTSDVHCLSHPHFTTLYRGSINTFLLTTKSDLGDIHSIRVWHNNEGRSPSWYLSRIKVENLFSRHIWLFICQKWLSVDTTLDRTFHVT >DENND5A_homSap_PLAT chr11: 9116949 RAB6 interacting 1 RUN-LH2-RUN pdb:3CWZ LIHILIVPSKKLGGSMFTANPWICISGELGETQIMQIPRNVLEMTFECQNLGKLTTVQIGHDNSGLYAKWLVEYVMVRNEITGHTYKFPCGRWLGKGMDDGSLER >DENND5B_homSap_PLAT chr12: 31426424 RAB6 interacting 2 RUN-LH2-RUN pdb:3CWZ PYRSVIIPIKKLSNAIITSNPWICVSGELGDTGVMQIPKNLLEMTFECQNLGKLTTVQIGHDNSGLLAKWLVDCVMVRNEITGHTYRFPCGRWLGKGIDDGSLER >ALOX5_homSap_PLAT chr10: 45189635 arachidonate 5-lipoxygenase NH2-LH2-lipox pdb:2P0M PSYTVTVATGSQWFAGTDDYIYLSLVGSAGCSEKHLLDKPFYNDFERGAVDSYDVTVDEELGEIQLVRIEKRKYWLNDDWYLKYITLKTPHGDYIEFPCYRWITGDVEVVLRDG >ALOX15_homSap_PLAT chr17: 4480963 arachidonate 15-lipoxygenase NH2-LH2-lipox pdb:2P0M GLYRIRVSTGASLYAGSNNQVQLWLVGQHGEAALGKRLWPARGKETELKVEVPEYLGPLLFVKLRKRHLLKDDAWFCNWISVQGPGAGDEVRFPCYRWVEGNGVLSLPEG >ALOX15_oryCun Oryctolagus cuniculus (rabbit) arachidonate 15-lipoxygenase 15S-LOX1 PDB:P0M GVYRVCVSTGASIYAGSKNKVELWLVGQHGEVELGSCLRPTRNKEEEFKVNVSKYLGSLLFVRLRKKHFLKEDAWFCNWISVQALGAAEDKYWFPCYRWVVGDGVQSLPVG >ALOX12_homSap_PLAT chr17: 6840108 arachidonate 12-lipoxygenase NH2-LH2-lipox pdb:2P0M GRYRIRVATGAWLFSGSYNRVQLWLVGTRGEAELELQLRPARGEEEEFDHDVAEDLGLLQFVRLRKHHWLVDDAWFCDRITVQGPGACAEVAFPCYRWVQGEDILSLPEG >ALOX15B_homSap_PLAT chr17 7,888,129 arachidonate 15-lipoxygenase NH2-LH2-lipox pdb:2P0M AEFRVRVSTGEAFGAGTWDKVSVSIVGTRGESPPLPLDNLGKEFTAGAEEDFQVTLPEDVGRVLLLRVHKAPPVLPLLGPLAPDAWFCRWFQLTPPRGGHLLFPCYQWLEGAGTLVLQEG >ALOX12B_homSap_PLAT chr17: 7916679 arachidonate 12R-lipoxygenase NH2-LH2-lipox pdb:2P0M ATYKVRVATGTDLLSGTRDSISLTIVGTQGESHKQLLNHFGRDFATGAVGQYTVQCPQDLGELIIIRLHKERYAFFPKDPWYCNYVQICAPNGRIYHFPAYQWMDGYETLALREA >ALOXE3_homSap_PLAT chr17: 7939943 arachidonate lipoxygenase 3 NH2-LH2-lipox pdb:2P0M AVYRLCVTTGPYLRAGTLDNISVTLVGTCGESPKQRLDRMGRDFAPGSVQKYKVRCTAELGELLLLRVHKERYAFFRKDSWYCSRICVTEPDGSVSHFPCYQWIEGYCTVELRPG >LIPC_homSap LIP-PLAT chr15:56511467 lipase C YHYQLKIQFINQTETPIQTTFTMSLLGTKEKMQKIPITLGKGIASNKTYSFLITLDVDIGELIMIKFKWENSAVWANVWDTVQTIIPWSTGPRHSGLVLKTIRVKAGETQQRMTFCSEN >LIPG_homSap LIP-PLAT chr18:45342425 endothelial lipase YHYQMKIHVFSYKNMGEIEPTFYVTLYGTNADSQTLPLEIVERIEQNATNTFLVYTEEDLGDLLKIQLTWEGASQSWYNLWKEFRSYLSQPRNPGRELNIRRIRVKSGETQRKLTFCTEDPEN >LPL_homSap LIP-PLAT chr8:19840862 lipoprotein lipase FHYQVKIHFSGTESETHTNQAFEISLYGTVAESENIPFTLPEVSTNKTYSFLIYTEVDIGELLMLKLKWKSDSYFSWSDWWSSPGFAIQKIRVKAGETQKKVIFCSREKVSHLQKGKAPAVFVKC >PNLIP_homSap LIP-PLAT chr10:118295418 pancreatic lipase WRYKVSVTLSGKKVTGHILVSLFGNKGNSKQYEIFKGTLKPDSTHSNEFDSDVDVGDLQMVKFIWYNNVINPTLPRVGASKIIVETNVGKQFNFCSPETVREEVLLTLTPC >PNLIPRP1_homSap LIP-PLAT chr10:118340480 pancreatic lipase-related 1 WRYGVSITLSGRTATGQIKVALFGNKGNTHQYSIFRGILKPGSTHSYEFDAKLDVGTIEKVKFLWNNNVINPTLPKVGATKITVQKGEEKTVYNFCSEDTVREDTLLTLTPC >PNLIPRP2_homSap LIP-PLAT chr10:118370455 pancreatic lipase-related 2 WRYKVSVTLSGKEKVNGYIRIALYGSNENSKQYEIFKGSLKPDASHTCAIDVDFNVGKIQKVKFLWNKRGINLSEPKLGASQITVQSGEDGTEYNFCSSDTVEENVLQSLYPC >PNLIPRP3_homSap LIP-PLAT chr10:118177414 pancreatic lipase-related protein 3 chr16:70520943 WRHKLSVKLSGSEVTQGTVFLRVGGAVRKTGEFAIVSGKLEPGMTYTKLIDADVNVGNITSVQFIWKKHLFEDSQNKLGAEMVINTSGKYGYKSTFCSQDIMGPNILQNLKPC >PLAT_cloPer Clostridium perfringens phospholipase toxin DQ183948 KELVAYISTSGEKDAGTDDYMYFGIKTKDGKTQEWEMDNPGNDFMTGSKDTYTFKLKDENLKIDDIQNMWIRKRKYTAFPDAYKPENIKVIANGKVVVDKDINEWISGNSTYNIK >ALOX_8R_pleHom Plexaura homomalla (coral) 8R-Lipoxygenase LOX-8R beta calcium membrane AIYNVEVETGDREHAGTDATITIRITGAKGRTDYLKLDKWFHNDFEAGSKEQYTVQGFDVGDIQLIELHSDGGGYWSGDPDWFVNRVIIISSTQDRVYSFPCFRWVIKDMVLFPGEA >LOXHD1_galGal_PLAT:01 KVYEVVTVTGDVRGAGTDANVFVTLFGEFGITPKTHLTSKSSTAFERSKTDVFRVKTNNVGQIKKIRIEHDNTGLNAGWFLDRVIVTDMNRPhLRFYFPCNNWLSKEDGDGLYVRDLI >LOXHD1_galGal_PLAT:02 NKYVVRVFTGEVSGSGTDADVFINIFGEKGDTGVRKLDNDKDNFEKGAEDKFTLDAPNLGRLRKINIGHNNKGGSAGWFLAKVIIEDIGNKCVYQFPVGRWFALDEDDGKIQRDIL >LOXHD1_galGal_PLAT:03 IVYNVAVVTGDIRGAGTNSKIHVILHGSKglKNSGTIFLEGGEFERARTDLFNVEIASLLSPLSRVTIGHDNCGVSSGWYCEKVVVYCPFTGIEQTFPCGKWLDEDEGDGLIERELY >LOXHD1_galGal_PLAT:04 NPWSLWIWTSDIKNAGTDATIFFQIYGDKGKSDEMKLDNNSDNFEAGQTDKFMIELPDLGTFYKLRIWHEKRNPFAGWHLDKVTLLKTLTKDKYSFNCGRWLDINEDDNEIVRELP >LOXHD1_galGal_PLAT:05 IKYRVTVCTGMVSGSGTDANVFVCLIGDQGDTGDRVLYKCinNVNKFEKGNADEFFVEAVTLKQVRRVRIGHDGKGGSSGWYLAKVIVREEGQPesEAVEFPCYRWLDKNEDDGQIVRELV >LOXHD1_galGal_PLAT:06 VSYHISVKTGDIPGASSDSKVFIKLYGEKADTSKETLLVSdndLGNYFERGRVDEFTIDTMDIGKINRILIGHDNVGLRSGWFLASVQITVPVQGRQYMFPCNRWLDKDEADGRVEVEVY >LOXHD1_galGal_PLAT:07 INYEVSVVTGDVRAAGTNAKVFMQIYGETGKTELIILENRSNNFERGATDIFEVEAADVGKIYKIRIGHDGKGIGDGWFLESVTLKRLATKmdetdkkkkkkkkkseeeeeeeetkveevmDVYTFVAHRWLAKDEGDKELVVELV >LOXHD1_galGal_PLAT:08 NTYEVHVLTGSVWGSGTDANVFLSIYGIErGDTGERQLKRSnNLNKFEKGQVDVFTIKAIDLGELKKLRIRHDNSGSSPSWFLERVEIVDLKESTTYYFPCQRWLAVEEDDGQIVRELV >LOXHD1_galGal_PLAT:09 TTYIVKVKTGDKKNAGTDANVFITLYGSKDDTGIVSLKASklNKNKFERGKIDEFTVESVDIGDLKKIKIGHDNAGNSNGWFLEWVEIDAPSLGQCLKFPCGRWLDKSEDDGAIERFIF >LOXHD1_galGal_PLAT:10 VPYEITVYTSDIFGAGTDADVFIVLYGSDgicTQQKSLCLNKReQRMYFERNSVNQFIVELEDVGDIIEKIRIGHNGGGLNSGWHLDRVAIRRLLPNgkgsETITFPCERWLAKSEDDGEIIRELV >LOXHD1_galGal_PLAT:12 IPYHITVTTGTEYDSSTDSRVFIIIMGPQkVRTERLWLDLPeGKDEFADGSVEKFSVWGLDVGEIKKVEVGHDGATPESCWLMEELTIVVPTKGVMYNFVCKCWLARDKGDGLTSRILN >LOXHD1_galGal_PLAT:13 ILYEVTVVTGDIESGGTDAGIFMTVFGSNGNTEEMQLDKNGDRFERGQEDSFIMEIADIAPLRKMRIRTDAKGTRPDWFLERIVMRNLTNQEVATFTYGDWLSKVKNAKGSLVCEM >LOXHD1_galGal_PLAT:14 TTYTIQVKTSDIGGAGTDANVSLILFGENGDSGTLALKESnKSNKFERNQMDEFNFPNMLSLGDLCKVRIWHDNKAY >LOXHD1_galGal_PLAT:15 KAYEIVTVTSNREDAETKENIWIILEGKLGRSKEFLMENSskKRRFERRGSTDTFQFSSKNLGDIAAICVGHCPKDgkkssakADVYWHVKEIIITEMELCNKYFFRCNGKIPLRYKRRDYKVFEC >LOXHD1_galGal_PLAT:16 VKYETIVVTGFEKGAGTDANVFITIFGLNGDSGKRALKQKFRNLFERGKTNRFYLETLD MGELKKVRIEHDNSGLAPGWLVERVEITNSATGVTTIFPCGKWLDENRGDGLTWRELF >LOXHD1_schMan_PLAT:01 Schistosoma mansoni (flatworm) Bilatera; Platyhelminthes; Trematoda XP_002576380 from contig NS_000200 TLYKVVLTTADVPGAGTSAQIYITLKGEWGSSTRQKLRKEKVPTRNLRFYFYPGSTNTFSVVSPDLGGLHSVFIEHDSLRKSDSWLLESVQVFHPLTKKRYMFMCNHWFSLYKEDGLIARELF >LOXHD1_schMan_PLAT:02 TKYSIVTVTGDQEGSGTNSKVYITIYGRTGITPRIELSQENKSTGKDILCAPFGRGTSTKFIVKAPNVGAITNIRIKQDESGNEPHWFLERVVVTDMSYPQWTYYFHLSCWLSSKYGDGKSCR >LOXHD1_schMan_PLAT:03 TEYKLTFYTSDQKDAGTTGEVYVKLYGEKDSSREIWVNSINQKNRRQPTSYHFTRGSTVEVYLPPCPQLGEITKLKVGHNRAGSSPSWFLNKVIVDDLRMNRVFEFPCYAWITK >LOXHD1_schMan_PLAT:04 IPLEIRLYTGDVPNAGTTAKVYLHQKTTSENKEDVYTTPYIWLEDGNYDRNGVTLFSIDLPVPKFISPLSQLVIGHDNTGHSPSWFFDKVQIYCPLNGVEQTFLYRKWLTSSKPGMKVEQILCEE >LOXHD1_schMan_PLAT:05 IPWEISVKTSPIANSYVTAAVSIIIFGSKDKTMKIQLDRSNLEVSSEWTKVSGNEVVEMFRPNVESKFRIYIKDIGIPCKLRIQHDNKGSNPNWHLQEIVLTNLRTHEQYEFYCNRWLSTREDDGSILREIPAK >LOXHD1_schMan_PLAT:06 YIIQIFTGNKPNSGTNANIFINIFGEKGDCGERWLGRSVNRNSELFQQNQMDEFVIEAVQLGSINKICIGHEERSPGYGWYLAKIVLTIKENPKYKLTFECYRWFDVGEDDGQIVRELFA >LOXHD1_schMan_PLAT:07 IAYNVTVLTGTCRNAGTVANVFVHLYGLQGESKDMQLKHKETEITKFEAGKSEEFILACGKLGEIKKIKIWHDSIAPHSGWFLDEVVVIDYFLGRRYVFHVNRWLAKNEADGLISLDLQPT >LOXHD1_schMan_PLAT:08 SYEVIVKTGGRKYAGTDSHVYITMFGINSESKEYHLANSKTHNNKFEQNHEDLFNLNAVGLGSLKKIRVRHTNTGIAPGWFLDYILIRESIEQKKMEYFFPCYQWLSATRLDGLVIREL >LOXHD1_schMan_PLAT:09 TIYHVRIHTGSQSNISSDANVQIQLYGDKDFTGNIRLYKAYYGKEDILVNKFQAGQIANFIVKAINIGNIKKIRIEHDTAVGSARWFVERVEVEARKLGLLWKFECNR >LOXHD1_schMan_PLAT:10 TNYQIKTFTSDISGAETTSSVYIQLYGNDSLPSSIRRLHQNNDSEQRFQRNKIDTFYVELEELQEPFSKLRIWHNDKGSSTDWHLNKVEIRKIKTNQYVFVTYIFPCNKWLSRNMDQAALERELIPS >LOXHD1_schMan_PLAT:11 NTYEVRITTGDKAYAGTDASVYITLFGENGDSGERKLTKSLTHRNKFERGQTDVFQLEIVDLGKINKVRIRHDNSGVNPSWYLSTIEVFNISKSQSVHGLKDQPNETHHLVKYIFNCEAWLSTEHDEKVLDR >LOXHD1_schMan_PLAT:12 IPYLITVITGSDRKAGTPGPVWISCVDKDKMNSEKFILCDCYNRTMLKRGTTRYFRFAGVKLNDLTEIQVGNDAPESPSMGWYIQSLYLSFPTIGKMYLFDCKEWLSTNRGNKK >LOXHD1_schMan_PLAT:13 ITYHLKITTANVKRAGTDCSINLQIFGTNGVTNCYILEKTSNRFAQGITDNISLEMEDVGKLLKMRIGHDNQGKNKHWNLSCVEVTVANTNQLYRFVYDDWLSLTYGKRKSLWADLPAM >LOXHD1_schMan_PLAT:14 TCLDIFVKTGNMPASSTDANVYVQLFGEYGDSGEILLKQTVSNQKPFQNNSIDHFKIPSILKLGNLARCRIWHDNKGSSPNWYCEWLEVKEVLIPGEKNLACNWKFAFNKWLSVSDDNKQLLRDAPCS >LOXHD1_schMan_PLAT:15 VYEVVIETGNLKDSGTTCDAWIILEGKHGRSPKLELVNQVGNPILQINQMNTFQLPSFPLGDLETIRLGIQERNINKQTNPNDIQSQKWFCEKVSIKDPVSKRTYIFTIHQWLS >LOXHD1_schMan_PLAT:16 VMYKVSIYTGTKGCANTDANIFITMFSTTPGLNSGRIALKRENNNLFDRKQLDEFYVESIDLESIQRIIIEHDNTGVSPDWYLDKVLITNQTNNQIHLFQCYQWISKKKGDCRLWKELLVS >LOXHD1_triAdh_PLAT:01 VAYQFGVKTGDKKGSDTDAVYIQVIGTKDKIPKKRLFKKQETEKTERGNLFKFDKSTVEKFAVQHRDIGDPVKLIVEHDGNEKRHGWFLEEITLTNIQSKKSWLFPCHKWLSKYEGDRKLCYE LKPLAKAGK >LOXHD1_triAdh_PLAT:02 AVYEVSVLTGDKRGAGTDANVSVTLFGKHTSSPKIQLLKSSKHKNPFERNNTDEFKIRTRDVGKLSKIRIEHDNAGFGPGWFLDKVIICNLEKPNVKYYCPCNQWLAKDVGDKSISRD LTAYTDPNAAPSA >LOXHD1_triAdh_PLAT:03 YVYIVHTFTGNKRGAGTDANVYAVIFGDSGDTGEKRLDNSKNNFEKSRKDTFKLSCSCVGKLERLRIRHDNTGLFAGWYLDKVVVEDPQEQQSYTFYCRRWLSKTEDDGEICRDLI VSASGDDGDDSVAPKG >LOXHD1_triAdh_PLAT:04 YPYHIHVTTSDVKNAGTDAEVYVVMHGEGKKSKELNSGKLVLANSEKKKNTFERAMTDIFHMECAEMLSPLTKLTVGHDNKGLAAGWHLDRaIVIDCPTTGIEQTFLCQQWLDRKAGDGLTERELVEA FDMRKTRRPK >LOXHD1_triAdh_PLAT:05 QLWFAWIWTSDIRGAGTDANVSMQIYGDKGKSQEIKLGNNTDNFEQATLDKFKLEIDQVGVPYKLRIGHDNSNAFPGWHLDKVKLENMNDKEQYLFNCNRWLSRSEEDNEIIRELPAS GPNCPNYPI >LOXHD1_triAdh_PLAT:06 VIYEVSVHTGNKMGGGTDANVFIKIYGELGDSGYRPLKSSKSHNNKFERNQVDVFHIEAVTLKALKKIKIGHDGNNPGAGWFLDKVVIKELNGEASNEFPCNRWLSKSEDDGQIVRELFLK SDTPLLKT >LOXHD1_triAdh_PLAT:07 TSYHISVKTGDVRNAGTDANVFIQIFGAKDDTGRVRLKQSLNTSNKFERNRIDKFIIEAAQIGKIEKIIIGHDGKGLGSGWFLDYIELDVPSVGRLYRFSCHQWFDSTEGDRKVERELYPS ECIKSAAK >LOXHD1_triAdh_PLAT:08 IPYQISVHTGDIRHAGTDSNVFAVIYGENGKTEELKLRNKSDNFERGQVDVFKVECEDVGKLRKLRIGHDSAGMGSAWFLDKVYVRRLPPKSGKKSKETDEREEETAKKDADEPEKLDENNYLFVANRWLSKEEGDRQTVIEIS PVGVDGALAE >LOXHD1_triAdh_PLAT:09 MTYTIRVITGNKFGCGTNANVFINMYGEEGDSGERQLKKSETHTDKFERNQEDVFKISCLSLGELKKIKIRHDNSGFRPAWFLDKVIIEVGESKYQFMCDRWLAKDEDDGQISRELLPQ SDEQSRAEAIGASKDLQKKVAS >LOXHD1_triAdh_PLAT:10 TTYNVSVTTGDIKGAGTDANVHIVLYGEKDDTGLIHLKNSTTHSNKFERNQEDRFVVEAIDIGELKKIKIGHDNKGGMAGWFLNKVEIDIPSLGRRLLFPCGRWIDKGKDDGALER ELYPLNEAEETYRPH >LOXHD1_triAdh_PLAT:11 IPYEVTVYTTDKRGASTGANVYVVIYGEENQTEQASLEPDKKRRKQYFKNGAIDKFVLELDDVGEEITKLRIGHDGKGWGAGWHLDKVEICRLLDGGKASKKFTFQCNRWLASDEDDGAIVRELVPS EIVEKSSKDGGQVKTKVTKPTDGLKV >LOXHD1_triAdh_PLAT:12 KPYTIHVFTGDVDGAGTNANVFLTIFGESGDSGERKLAKSDTHYDKFERNQEDIFHIEAADLGRLFKVKIRHDNTGSLFSPAWFLNRIEIVDDENEETTAFPCERWLAKKKDDGKIDRTLFVK >LOXHD1_triAdh_PLAT:13 VPYNIKVTVGEESSKNFQETLHLELFGQIEEEKSGPIELSPEKKSDKTFYPGKITTFYVSAAEVNIIEKIQVSHNSYMPDSGIYLKEIEVDVPTIGNKYIFPCNRWLAKDKDDSKTSRIFTAA >LOXHD1_triAdh_PLAT:14 KPYELTIHTGDVQNAGTDSNIFVILFGTKGRTPEISLEKNEDRFERAKVDIIPLELDDVGTIKKIRIGHDGKGSRTDWYLEKASIQRMDTLDMYMFRANQWFSKKIDDKKLVREI PAETSKEGATTIKK >LOXHD1_triAdh_PLAT:15 INYVLSTHTSDKRGSGTDANVFVIIFGENGDSGEIALKKSETNWNKFEKGQTDVFLINDRLSLGRLQKLRIWHDNAGFGASWHLASVDIVDESTGVKYTFPCDKWLSKSNGDKLILREL PCAETAGNTAASKATKQESKSGK >LOXHD1_triAdh_PLAT:16 AEYEIAFTTGTEKRAGTNQDVAIVLKGKSEKSREFLIENNEDKKYFSKGKTNKFTYTCKPLGDITKAIVSHRESAIGEEPDSKNSSWYLKVVTVVHKASGTTYKFPCNKWIDLDDDEENNSSVTLKC KSADVAASSKAKPVELKP >LOXHD1_triAdh_PLAT:17 VKYEVTVVTGDEKGAGTDANVSVILYGDNGDTGPRPLKKKFVNLFERNQHDKFTIEALDLGKLTKLHIEHDNKGWGASWLLDRVEVHNVDSNETIIFPCKQWLDKKKGDGQIAKDLLPES* >LOXHD1_monBre_PLAT:01 Monosiga brevicollis (monosiga) GHASLEIFTSKRSNPVSKSEKYITVVGTKGRSDAIQLAAPGVHFRAGNKDVFQVNLTGIGKPTKVILENTGTKRTDGWCVSKVVLVKKTDKGTRRYRFSGPVWLSKHHDEMKLKR >LOXHD1_monBre_PLAT:02 Monosiga brevicollis (monosiga) NGYRIDCYTGDVANAGTDAVATIQLFGSKGQSPMVELRRSDGQAFQRARVATFTLDDLANLGKLKKLVISHNGHGMASGWFLDKIIVTSLSSNKATVFPCDAWLDRKNGRSKELVAR >LOXHD1_monBre_PLAT:03 Monosiga brevicollis (monosiga) TTFTIRVMTGDRRGAGTDANVQCTLFGEDGESGPHTLNTSRNDFRRGHTDVFAVSSRKIGTLKRLRIWHDNGGAGPAWFLDAVEVVDEASGQTYRFECNRWLAKDEDDGQISRELTCN >LOXHD1_monBre_PLAT:04 Monosiga brevicollis (monosiga) KSYKLTIFTGDKRNAGTSANVFCKLVGERGASDNVILENSSKNFQRDRTDIFTVEASDLGSLRHIVLGHDNHGMGAGWYVERFSLEVPSEGKLYNVDVKQWFATDMSDGAIERTFRLD >LOXHD1_monBre_PLAT:05 Monosiga brevicollis (monosiga) LDWKCTIYTSDVANAGTDANVFMQVYGKKGKTDVVPLKNKSDTFERGQTDELRVQLINVGSLRKLRVWHDNKGMASGWHLDRIVLSRDGEEYIFPCAEWLAVSEGDKEIVRELPAT >LOXHD1_monBre_PLAT:06 Monosiga brevicollis (monosiga) VEYTVRVATGHARFAGTNADVFVMLTGELGDSGKRALLRSQTNRNKFERGKEDVFTVAAVDLGKLTSVTVGHNNAGTSAGWFLDKIVVLDPRRGEEEEFPCHRWLAVDADDGQIERELVPK >LOXHD1_monBre_PLAT:07 Monosiga brevicollis (monosiga) TTYIVKIKTGDVRHAGTDANVFVQLFGKTGESTQLKLRNSETYSDAFERNKMDIFKFELLDLGDLSRILVGHDNKGMGAAWFLDYVEVEVPSIRTRWKFPCSRWFSKSQDDGLTER >LOXHD1_monBre_PLAT:08 Monosiga brevicollis (monosiga) APYLFRFYTSDVAFAGTDANVSVVLYGDEGKTEELVVNNQSDNFERGKADDFKLACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQARIYTFEHNDWLRKGTKAKPFMVEL >LOXHD1_monBre_PLAT:09 Monosiga brevicollis (monosiga) RTYRVKVHTGDQKGAGTDANVYVNLHGSLGDSGDRHLKNSLTHTNKFQRKTVDEFDIDAVTLGDINKVKVWHDNAGLGAAWYLEKIEVVDTADDKTYIFPCAQWFAKSMGDGQIAREL >LOXHD1_monBre_PLAT:10 Monosiga brevicollis (monosiga) FKYRISVHTSDVKHAGTDANVDIVLYGEKGDTGKIRLAKSETHRDMWERGNCDVFTVSAIELGDLKRVDIMHDGKGVGSGWHLNKVVVDAPQAGKTWTFMCDAWLDKATDDGTMAK >LOXHD1_monBre_PLAT:11 Monosiga brevicollis (monosiga) VPYEIIIKTSDVRNAGTDANVFIDLYGRDQEERDLTAHHEFKDAVKAHFERNLEDRFNVELPDVGSIYKIRLGHDGKGMSSSWHVASVVVINQRTHERFEFPCDAWLSKDKDDKKLVREFAVG >LOXHD1_monBre_PLAT:12 Monosiga brevicollis (monosiga) AIYKIHVFTGDIKHAGTDANIYVQIFGDTGDSGEIKLEKSETYRDKFERGHEDIFTHRCLDLGPLRKIKVRSDGKGLMGGDWYLDRVEVHQENDLSEPPVRFVCQDWFKRGKQEGDTLEREITAQ >LOXHD1_monBre_PLAT:13 Monosiga brevicollis (monosiga) LEYKVTVYTGTDTQAGTTANVWLQLFGEKDAPLTPAPPSPSKLSRSGSLFGRRRRASSDAQSVSSSLSAGASGPRETSTGRLQLNNAPADLQSGAKTTFTVTGLDVGELVGLEIGHDDERDKWYLEQVVVEVPKSATHAARRYEFKAGVWLAARADGSTSGSAK >LOXHD1_monBre_PLAT:14 Monosiga brevicollis (monosiga) LEYTLKVYTASADGAGCTAVPQVQLFGDKHTTEALPLRAGGDILPASVVETQHRVPDLGALLKVRLMVPRGSSWTVEKVEFGRAGQTPITFVGSDGSAVTLGAERLS >LOXHD1_monBre_PLAT:15 Monosiga brevicollis (monosiga) TSYRVYVTTADERGTGTDANVSIILYGAMGDSGEHSLTKSETFDDPFERGNTDVFTLEVPDLGELQRARIWHDGKGMFSSWKLDKIVVVVEATQSRYELPCGQWLSKNKGDKQLTRDLAVASKR >LOXHD1_monBre_PLAT:16 Monosiga brevicollis (monosiga) GTYKLEVATDSRAGGGCKGPVRIMLLDAEKNQLPLTLEPPGGEFAPGSVEHLVFDNVMLLGPLTELRIRRKPSGASSRRGAEDDDEDDNEASGASSQSGSAVSPWHLEHIIVKHLQSGQSFVFKGPSKGLSRSRAKLSVHTEA >LOXHD1_monBre_PLAT:17 Monosiga brevicollis (monosiga) AQYEVAVTTGTERGAGTDSNVFVTLFGKNGDSGERALAKSKTFRNMFESGNTDVFDVECQDLGELTKIEVKSDLKGFGAAWQLDKIKVTRTGSQNSWQFKCDQWFDKKQGAEHTFSVA
Reference PLAT:07 domains
>PLAT:07_homSap Homo sapiens (human) 2 LVHYEVEIWTGDVGGAGTSARVYMQIYGEKGKTEVLFLSSRSKVFERASKDTFQ 0 0 LEAADVGEVYKLRLGHTGEGFGPSWFVDTVWLRHLVVREVDLTPEEEARKKKEKDKLRQLLKKERLKAKLQRKKKKRKGSDEEDEGEEEESSSSEESSSEEEEMEEEEEEEE FGPGMQEVIEQHKFEAHRWLARGKEDNELVVELVPAGKPGPE 1 >PLAT:07_monDom Monodelphis domestica (opossum) 2 LIHYEVEIVTGDMGNASTSARVYMQIYGEEGKTEVLFLKSRSKVFQRGNTDTFQ 0 0 LQEAEVTPEEEARKKKEMDKLRQLMRKERLKAKEEQKRKRKLAKGSDVEDEEESSSEESSEESEEETEIEEEDEE FQEVVDVYRFHAGRWLATDEEDKDLDMELAPSGRSGPQR 1 >PLAT:07_ornAna Ornithorhynchus anatinus (platypus) 2 LIHYEVEIVTGDMGYAGTNARVYMQIYGELGKTEVLHLTSRTNVFEQGATDTFQ 0 0 LEATDVGEIYKVRLGHSGEGFGSGWFIESLVLKRLVLKEVEPNPEEEKRKAKERERAREQRRKERLKAKQQRKKKKKMKKSSDDEDSEAEDSEEEEGSSEEESSSSSSEEEVEEEFGPGIKEVIDVYKFEAHRWLARDEDDKELIVELEPANRPGPE 1 >PLAT:07_galGal Gallus gallus (chicken) 2 LINYEVSVVTGDVRAAGTNAKVFMQIYGETGKTELIILENRSNNFERGATDIFE 0 0 VEAADVGKIYKIRIGHDGKGIGDGWFLESVTLKRLATKMDETDKKKKKKKKKSEEEEEEEETKVEEVMDVYTFVAHRWLAKDEGDKELVVELVPDGESELE 1 >PLAT:07_taeGut Taeniopygia guttata (finch) 2 LINYEVSVVTGDVRAAGTNAKVFMQIYGETGKTELIILENRSNNFERGATDIFK 0 0 REAADVGKIYKIRIGHDGTGIGDGWFLESVTLKRLATKTEGSDKKKKKKKKSEEEETKEEEGMDVYTFVAHRWLAKDEGDKELVVELVPDGESDLE 1 >PLAT:07_anoCar Anolis carolinensis (lizard) 2 LINYEVCVATGDVRNAGTNANVFMQIYGELGKTELLILKNRSNNYERGASETFR 0 0 VEAVDVGKVYKIRIGHDGKGFGDGWFLDSVVVKKLPTKVPKKKKKKKKKKTPEEEEAEEGPGIMEVYNFTPCRWLASDEEDKELVVELVPDEGSELE 1 >PLAT:07_oryLat Oryzias latipes (medaka) 2 VINYEIAVVTGDVRAGGTNASVFCQIYGEEGKTEVLNLKSRSNNFERGTTEIFK 0 0 IEALDVGKIYKIRIYHDGSGIGDGWFLETVDIKRLTMALVQVEVKKEEAPKKDKKKDKKKKKKEEEEVEIIEEMQEVVETFTFTCNRWLARDEEDGEIVVELLTEENEDLE 1 >PLAT:07_gasAcu Gasterosteus aculeatus (stickleback) 2 VINYEVTVVTGDVTFAGTNARVIIQIYGDKGKTEVIILESRSNNYERNTTEIFK 0 0 IEAKDVGKIFKIRIGHDGSGIGSGWFLETVDVKRLILALVPKEKKKEDKKKKKKKKEDVDEEGGEEMQEVVLTYSFPCSRWLAGGEEDGELVVELLPDDAKELE 1 >PLAT:07_takRub Takifugu rubripes (fugu) 2 VINYEVTVVTGDVTFAGTNAKVFVQIYGDKGKTEVIMLESRSNNYERNAMEIFK 0 0 IEAKDVGKIFKIRIGHDGLGIGSGWFLEKVYVKHLIMALVPRENKKDDKKKKKKKKKDKEDEEEVGGEEMQEVVVTYHFPCSRWLASGEDDDDLVVELLPEDAEELE 1 >PLAT:07_petMar Petromyzon marinus (lamprey) 2 GINYQVSVHTGNVRSAGTNANVFIQLYGDMGKTEVHNLRNRSDNFERASNDIFK 0 0 VEAMDVGKVVKLRVGHDNSGMGSGWFLDSIVIRRLRQSSPHRPQPVDAEEDEDEEDDEEAEDEDSEEVQTYTFPCKRWLARDEDDGEIVRELLPQDCAEME 1 >PLAT:07_sacKow Saccoglossus kowalevskii (acornworm) Bilateria|Deuterostomia|Hemichordata 2 GIPYEITVITGDKSGAGTDAQVFIRMYGLKGKTDEFILNNRTDNFERGMTDKFK 0 0 IEAADVAMLTKIRIGHDNSGRSAGWYLERVIIERFPPKRKMKRKRSGTPRRRGEYDEEDYDDIPETNVVNFVCNRWFAKDEEDHQIVRELLPTDEEALKGH 1 >PLAT:07_strPur Strongylocentrotus purpuratus (sea_urchin) 2 GIPYEITVITGDKSGAGTDANVFLTMYGDDGAKTEEFSLRNRTDNFEKNMTDKFK 0 >PLAT:07_triAdh Trichoplax adhaerens (placozoan) Eukaryota|Metazoa|Placozoa 2 KIPYQISVHTGDIRHAGTDSNVFAVIYGENGKTEELKLRNKSDNFERGQVDVFK 0 0 VECEDVGKLRKLRIGHDSAGMGSAWFLDK 00 VYVRRLPPKSGKKSKETDEREEETAKKDADEPEKLDENNYLFVANRWLSKEEGDRQTVIEISPVGVDGALA 1 >PLAT:07_monBre Monosiga brevicollis (choanoflagellate) Eukaryota|Choanoflagellida 2 PMEEDVSAPYLFRFYTSDVAFAGTDANVSVVLYGDEGKTEELVVNNQSDNFERGKADDFK 0 0 LACKPVGRPSKIRLSAHGGGMSADWHLEKIEVHELGQARIYTFEHNDWLRKGTKAKPFMVELPLRRIETVDDNGREVVEELALDANKRTYR 1
See also: Usher: USH2A | Usher: CDH23 | RBP3 (IRBP) | RPE65 | Transducins